Tau蛋白病是一类以异常磷酸化的tau蛋白聚集为病理特征的神经退行性疾病,包括阿尔茨海默病(Alzheimer’s disease, AD)、皮质基底节变性(cortical baseal degeneration, CBD)、进行性核上性麻痹(progressive supranuclear paralysis, PSP)、亨廷顿病(Huntington disease, HD)、皮克病(Pick disease)和唐氏综合症(Down’syndrome)等多种疾病
[1]。Tau蛋白是一种微管相关蛋白,对于维持细胞骨架稳定和轴突胞浆运输具有重要作用。病理条件下,tau蛋白异常磷酸化可导致微管相关蛋白功能失调,细胞骨架崩解,轴突胞浆运输障碍,最终引起神经元变性坏死和突触丢失
[2]。但目前tau蛋白病的病理机制尚不完全清楚。越来越多的研究提示自噬与tau蛋白病发生发展密切相关。Tau蛋白作为长寿命蛋白,在正常生理情况下可以通过泛素蛋白酶途径和自噬途径来降解。在最常见的tau蛋白病AD病理情况下,脑内自噬水平明显异常,提示自噬可能参与tau蛋白病
[3-4]。研究表明,p62作为选择性自噬的重要受体之一,能够通过溶酶体介导自噬来维持细胞内环境稳态
[5],同时参与细胞信号传导和蛋白的运输、聚集和降解。特别是,p62存在于神经纤维缠结(neurofibrillary tangles, NFTs)中
[6],进一步提示p62作为自噬重要组成部分和tau蛋白病关系密切。但tau蛋白病中p62的变化情况以及p62和tau蛋白磷酸化之间关系尚无报道。为此,本研究以含人tau基因P301L突变的转基因小鼠为模型,通过检测p62和tau蛋白过度磷酸化的关系,来初步探讨p62在tau蛋白病发生中的作用。
1 材料与方法
1.1 实验动物
雄性含人P301L突变tau转基因小鼠(P301L转基因小鼠)由澳大利亚昆士兰大学Juergen Goetz教授提供。该小鼠携带人致病型Tau P301L突变基因。采用3月龄和9月龄雄性P301L转基因小鼠各4只,以同窝同龄野生型雄性小鼠作为对照。
1.2 主要试剂
兔抗tau(Phospho-Thr231)抗体(Signalway Antibody);兔抗SQSTM1/p62抗体(Abcam)。
1.3 方法
组织和切片制备:小鼠予以过量6%水合氯醛腹腔深度麻醉,经心脏灌注生理盐水冲净血液,肝脏和肾组织转白后解剖取大脑,置于4%多聚甲醛中4 ℃固定48 h,随后再将脑组织置于30%蔗糖溶液中。待标本沉底后,于冰冻切片机下行脑组织冠状位切片,厚度为35 μm。脑片在50%乙醇溶液中洗净,暂存于4 ℃ 0.01 M PBS 溶液中备用。
Western blotting:将脑匀浆直接上样在8%~12% 的SDS-PAGE中电泳,采用兔抗tau(Phospho-Thr231)抗体(1:2000),兔抗SQSTM1/p62抗体(1:2000),同时检测β-actin作为参照,用Quantity one 4.0软件分析各条带的光密度值,最终蛋白定量结果以“实验条带光密度值/β-actin条带光密度值”表示。
免疫组化:分别用鼠抗tau(Phospho-Thr231)和兔抗SQSTM1/p62抗体检测异常磷酸化tau蛋白和p62的表达。脑片在4 ℃冰箱中一抗孵育过夜后,加入生物素化二抗37 ℃孵育1 h,漂洗后用ABC试剂盒(vector laboratories)以DAB为底物显色10~20 s;常规贴片、脱水;以中性树脂封片。在光强度和聚光器设置保持不变的条件下摄片。用Image-Pro Plus (IPP6.0)软件定量分析转基因小鼠脑内异常磷酸化tau蛋白和p62表达情况,最终蛋白定量结果以平均光密度值“IOD sum/Area sum”( Mean IOD)表示。
1.4 统计学分析
计量数据以平均数±标准误表示,采用SPSS18.0 统计软件对P301L转基因和野生型小鼠间磷酸化tau蛋白和p62表达水平进行单因素方差分析。
2 结果
2.1 脑组织tau蛋白异常磷酸化和p62蛋白水平的比较
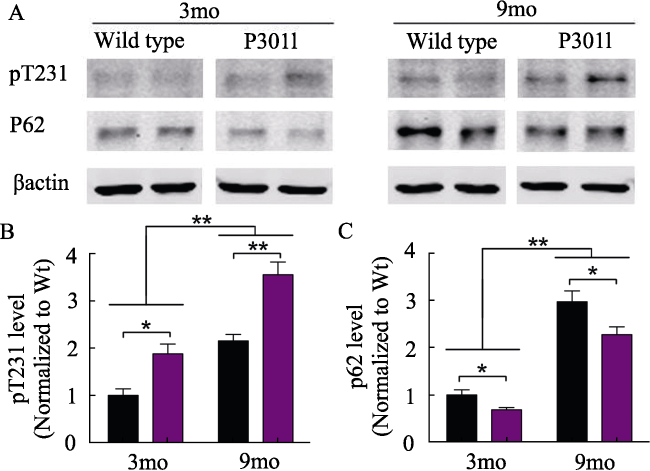
Western blotting 结果表明P301L转基因和野生型小鼠脑内tau蛋白磷酸化和p62蛋白水平均随着年龄的增长而增加(P< 0.01); P301L转基因小鼠tau蛋白磷酸化水平显著高于同龄野生型小鼠(P< 0.05),而p62水平低于同龄野生型小鼠(P < 0.05,图1)。
图1 Western blotting检测不同月龄小鼠脑组织内pT231- tau和p62蛋白表达水平。A: 3月和9月龄野生型小鼠(Wide type, Wt)和P301L转基因小鼠脑组织中pT231-tau和p62表达水平Western blotting检测结果;B和C:半定量分析比较。*: P< 0. 05; **: P< 0. 01。 |
Full size|PPT slide
2.2 脑组织切片海马tau蛋白磷酸化和p62表达水平的比较
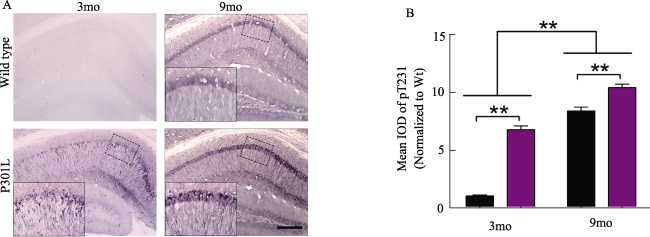
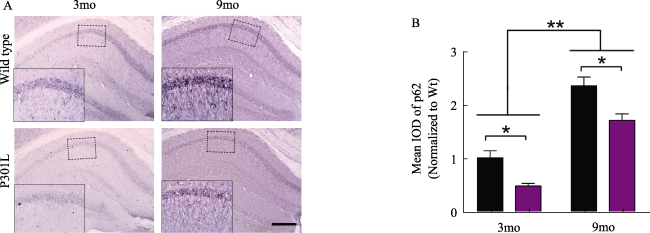
免疫组化染色结果表明在P301L转基因小鼠海马神经元大量表达pT231-tau,随年龄的增长,阳性神经元数量明显增多,着色加深。p62在小鼠海马区域表达随年龄增长而增加,P301L转基因小鼠海马p62的表达低于同龄野生型小鼠(图2、图3)。
图2 小鼠海马pT231-tau蛋白免疫组化(标尺 400 μm,放大倍数100倍)。A: 3月和9月龄野生型小鼠(Wide type)和P301L转基因小鼠脑片海马区域pT231-tau免疫组化染色观察;左下角插图为虚线框部分的放大图(放大倍数400倍)。B:定量分析比较。**: P< 0. 01。 |
Full size|PPT slide
图3 小鼠海马p62免疫组化(标尺= 400 μm,放大倍数100倍)。A: 3月和9月龄野生型小鼠(Wide type)和P301L转基因小鼠脑片海马区域p62免疫组化染色观察;左下角插图为虚线框部分的放大图(放大倍数400倍)。B:定量分析比较。*: P< 0. 05; **: P< 0. 01。 |
Full size|PPT slide
3 讨论
本研究通过检测p62在不同月龄雄性转基因小鼠及同窝野生型小鼠脑中的表达发现,p62和异常磷酸化tau蛋白水平在各组小鼠脑内均与年龄呈正相关,进一步表明p62是一个与老化相关联的蛋白分子。但与同月龄(3月龄和9月龄)野生型小鼠相比,P301L转基因小鼠脑内p62表达水平显著降低,尤其表现在海马,提示在tau病理进程中,p62蛋白可能发挥重要作用,在多个方面影响tau蛋白的异常磷酸化和神经纤维缠结的形成,如自噬水平的过度激活、蛋白修饰异常等。
tau蛋白异常磷酸化在tau蛋白病中发挥神经毒性作用,可导致脑内神经元变性坏死和突触的丢失等
[7]。Tau蛋白病的发病机理目前尚不明确,可能与基因变异、表观遗传学、蛋白质异常修饰和氧化应激异常等有关
[8]。研究表明,自噬与tau蛋白病的发生发展密切相关。正常自噬水平可以降解异常磷酸化的tau蛋白,而自噬功能障碍能够促进tau蛋白的异常聚集
[9]。
SQSTM1/p62是一种多结构域蛋白,结合入核出核信号穿梭于细胞核和细胞质间,不仅是选择性自噬的受体,也参与多种细胞信号传导、蛋白修饰以及抗氧化应激等,p62缺乏会加速老化导致老龄化相关的病理改变
[5,10]。在AD患者额叶皮层中,p62的基因和蛋白表达水平显著降低
[11]。在过度表达类淀粉样蛋白(amyloid-beta, Aβ)的AD小鼠模型中,p62蛋白表达也显著减少
[11]。而在p62敲除的小鼠脑内,tau蛋白磷酸化、神经纤维缠结形成、记忆相关的突触丢失和神经元的凋亡更为严重,且呈年龄相关性
[12]。与上述研究相一致的是,本研究发现tau蛋白病转基因小鼠模型中,p62蛋白表达显著下调。
在P301L突变的tau蛋白病模型小鼠脑内,尤其在海马、皮层等区域
[13],tau蛋白过度磷酸化,导致神经元丢失和突触损害,引起小鼠年龄相关的行为学损害,如记忆提取障碍、多度自发活动等
[14]。海马区域是影响P301L小鼠行为学的主要脑区之一。P301L小鼠海马区域p62表达量减少,提示p62的表达可能参与调节tau模型小鼠的行为学改变。
综上所述,本研究发现tau蛋白病转基因小鼠脑内p62表达异常,提示自噬异常参与了tau蛋白病的发生发展。


{kind=link}
{kind=link}
{kind=link}