
2022年第三期

胆汁酸稳态失衡与阿尔茨海默病研究新进展
日期:2023-11-22
阿尔茨海默病(Alzheimer's disease,AD)是一种起病隐匿、持续进展的中枢神经变性疾病,以进行性认知功能障碍为主要临床表现。目前诸多研究发现胆汁酸稳态失衡和信号转导异常与AD致病机制相关,胆汁酸谱有望作为该疾病的潜在生物标志物和治疗靶点。本文系统梳理了胆汁酸谱代谢变化特征在AD发病过程中可能发挥的作用以及对相关的胆汁酸潜在生物标志物进行总结,为疾病预防和治疗干预提供新思路。
Abstract
Alzheimer's disease(AD) is a insidious, rapidly progressive central neurodegenerative disorder, with progressive cognitive dysfunction as the main clinical manifestation. Currently, the imbalance of bile acid homeostasis and abnormal signal transduction are found to be related to the pathogenesis of AD. Therefore, bile acid spectrum is expected to be a potential biomarker and therapeutic target of AD. This study systematically summarized the role of metabolic changes of bile acid profile in the pathogenesis of AD and discussed its potential biomarkers, which provide the new avenue for prevent and cure AD.
关键词
Key words
Alzheimer's disease / Bile acid / Gut microbiome / Metabolic disorders
引用本文
李悦 , 豆小文 , 纪翔 , 李瑞 , 朱飞奇 , 张秀明. 胆汁酸稳态失衡与阿尔茨海默病研究新进展. 阿尔茨海默病及相关病杂志. 2022, 5(3): 242-247 https://doi.org/10.3969/j.issn.2096-5516.2022.03.015
LI Yue , DOU Xiaowen , JI Xiang , LI Rui , ZHU Feiqi , ZHANG Xiuming. Recent advances in study of bile acid homeostasis imbalance and Alzheimer's disease. Chinese Journal of Alzheimer's Disease and Related Disorders. 2022, 5(3): 242-247 https://doi.org/10.3969/j.issn.2096-5516.2022.03.015
阿尔茨海默病(AD)是一种进行性中枢神经变性疾病,临床潜伏期长,目前AD诊断主要依赖于临床认知和行为评估、实验室和影像学检查,而AD患者的病理学改变可开始于症状出现前的数年或数十年,大多情况下开始检查时神经损伤已不可逆转,难以实现早期诊断分型及治疗,因此AD早期的发现和治疗是现代医学的一大难题。探索该疾病代谢途径及相关标志物,对疾病的预防和治疗具有重要意义。
胆汁酸(Bile acids,BAs)是胆固醇在人体与肠道微生物共同代谢产生的最终代谢产物,具有调节和信号转导功能,在胆固醇代谢、脂质消化、肠道微生物的相互作用与调控等发挥着重要作用。近期临床研究显示几乎所有在肠肝循环中的BAs在脑组织中都可以检测到,提示人体中可能存在贯穿与神经变性病变机制相关的肠道菌群-肝-脑的胆汁酸代谢轴[1-2],因此BAs的肠道菌群转化可能与AD发生发展密切相关,BAs谱可能作为AD的潜在生物标志物。诸多实验数据和临床研究证实BAs参与AD的发病机制[2⇓-4],血清BAs谱的水平与大脑结构和功能的改变显著相关[5]。本文对相关文献进行综述,探究BAs谱代谢标志物和阿尔茨海默病之间的关联性,及其作为临床诊断标志物的可能性。
1 胆汁酸代谢与肠道微生物作用
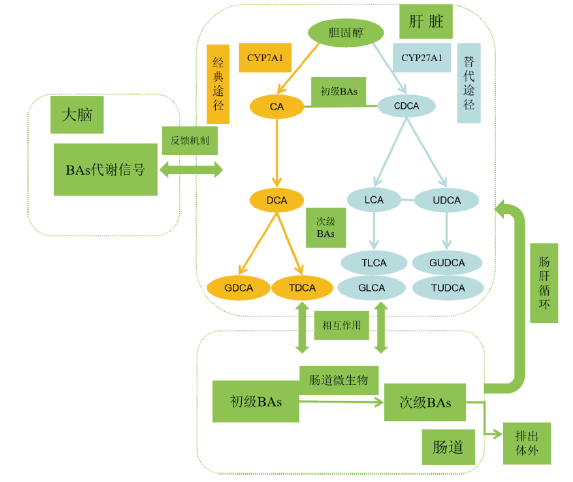
BAs合成受肝脏中的胆固醇7α-羟化酶(CYP7A1基因)调控,初级BAs通过肠道细菌中的酶活性转化为次级BAs,而次级BAs刺激肝脏进而抑制BAs合成,从而维持BAs稳态(见图1)。人体中BAs 具有清除胆固醇的作用,在肠道中作为清洁剂乳化脂肪,被肠道细菌代谢,帮助机体消化和吸收营养物质。95%以上BAs参与消化后在回肠末端重新吸收进入“肠肝循环”[6]。此外,BAs是重要的信号分子参与调节多种代谢过程,显著影响胃肠道运动、感觉、分泌功能以及肠道屏障通透性和炎症反应的调节[7]。
{kind=link}
肠道微生物群在病原体防御、免疫和营养收获中起着重要作用,诸多证据表明肠道微生物群、肠上皮屏障、肠神经免疫网络动态相互作用的变化可能是神经退行性过程的起点[8]。肠道微生物群组成改变导致肠道屏障通透性增加,损害血脑屏障,促进神经炎症,是可能引起神经退行性变的发病机制之一。该过程中BAs是调节肠道微生态失衡的重要指标,可以通过上调小肠固有免疫应答基因改变肠道菌群多样性[9]。疏水性胆汁酸如脱氧胆酸(Deoxycholic acid,DCA)等对细菌的生长可产生直接的抑制作用,对肠道菌群的结构组成发挥重要的调节作用[10];Bas还可激活法尼醇受体(Farnesoid X receptor,FXR)和G蛋白偶联胆汁酸受体(G-protein-coupled receptor for bile acids,TGR5)发挥信号传导作用,抑制微生物群过度生长和组成,保护肝脏和肠道免受炎症影响。反过来,肠道微生物群同样可以调节肠道生物转化,参与BAs合成和进一步修饰,改变BAs组成,并参与调节FXR和TGR5信号传导过程 [11⇓-13]。综上所述,胆汁酸与肠道微生物相互作用和相互影响,两者间的调节失衡参与多种疾病如AD、炎症性肠病和非酒精性脂肪性肝病等的发病过程[2,14-15]。
大脑通过反馈机制调节BAs合成和运输进入肝循环。图1中橙色代表经典途径,蓝色代表替代途径。经典途径由CYP7A1启动负责大部分BAs的合成,并通过CYP8B1和CYP27A1的后续作用产生CA和CDCA。“替代”途径由CYP27A1启动,并通过CYP7B1产生CDCA,替代途径也可以发生在肝脏以外的其他组织。在肝细胞中,大多数次级BAs与甘氨酸或牛磺酸结合分泌进入胆汁。BAs在小肠肠道细菌中的微生物酶作用下形成次级BAs。在回肠末端,大多数未结合BAs被重新吸收进入肠上皮细胞通过基底外侧BA转运蛋白和有机溶质转运蛋白亚基α分泌进入肠肝循环,小部分排出体外。CA:胆酸,CDCA:鹅脱氧胆酸,DCA:脱氧胆酸,LCA:石胆酸,UDCA:熊脱氧胆酸,GDCA:甘氨脱氧胆酸,TDCA:牛磺脱氧胆酸,TLCA:牛磺石胆酸,GLCA:甘氨石胆酸,GUDCA:甘氨熊脱氧胆酸,TUDCA:牛磺酸脱氧胆酸。
2 肠道微生物群-胆汁酸-脑轴与阿尔茨海默病
研究已证实[1⇓⇓⇓-5],AD发病及进展可能存在BAs谱稳态失衡,AD与肠道微生物群-BAs-脑轴的代谢紊乱密切相关(见图1)。有研究通过43名AD患者和43名对照组的粪便样本对比分析肠道微生物的组成,显示AD患者体内梭状芽胞杆菌和双歧杆菌下调[16]。Vogt NM等比较25名AD患者和25名对照组的粪便样本肠道微生物种群[17],前者厚壁菌减少,拟杆菌增加,整体微生物丰富度和多样性降低,比较9名AD组与31名非AD组的微生物群发现细菌丰度与脑脊液标志物p-tau和p-tau/Aβ之间呈正相关,AD患者在YKL-40脑脊液标志物与拟杆菌、浊度杆菌和SMB53(梭菌科)之间存在显着关联。而Jiang Y等采用高通量测序技术考察12例轻度AD患者治疗前后肠道菌群差异[18],轻度AD患者治疗后厚壁菌、变形杆菌、放线菌和梭菌丰度降低,拟杆菌丰度增加。肠道微生物群通过胆汁盐水解酶(BSHs)催化启动BAs代谢[19],肠腔内BAs水平降低与外膜中含有脂多糖(Lipopolysaccharide,LPS)的革兰氏阴性细菌呈负相关,而BAs浓度增加有利于革兰氏阳性细菌(如厚壁菌门)生长,它们通过7α-羟基化将初级BAs转化为毒性更强的次级BAs[20] 。BAs的组成和含量是调节人类肠道微生物群落结构的重要因素,其在调节肠道微生物组方面起着重要作用,具有神经活性的BAs已经成为肠道微生物群和大脑之间的直接通信通道[21]。BAs稳态失衡影响微生物种群变化,肠道菌群不仅在AD早期发生变化,而且影响AD的全程,AD的发生发展与肠道菌群结构的异常变化密切相关。
BAs代表与认知相关的肠-肝-脑轴的组成部分,免疫和肠道微生物失调导致细胞毒性的DCA及衍生物等次级BAs的蓄积,提示 AD患者肠道生态失调可能是在大肠和小肠中由CA 和CDCA 进行7α-脱羟基化的厌氧菌定植增强引起。由于BAs可以穿透血脑屏障并在大脑中积聚,导致其受体和靶标介导的代谢功能受损,为进一步研究肠道生态失调和肠-肝-脑轴在AD发病机制中的可能作用提供了依据。调节肠道微生物群组成可能降低AD发病风险,延缓其进展[22]。有研究表明AD患者肠道微生物群的多样性相较于正常人群降低,通过益生菌治疗可以减轻AD的精神和组织病理学表现,这是由于肠道微生物群的调节对神经元通路产生有益作用,从而延缓AD的疾病进展,为AD的治疗提供一种新的选择[23-24]。AD相关菌群组成特征的变化也将有助于更深入、更系统地研究宿主与肠道菌群在代谢和免疫中的相互作用,揭示肠道菌群在AD发生发展中的作用,对AD相关菌群结构模式的分析有助于了解AD的分子病理基础,为AD创新药物的开发和设计提供理论依据。此外,肠道微生物群还参与免疫、神经内分泌和神经通路调节,进而影响小胶质细胞的成熟和功能[25⇓-27]。肠道细胞损伤和微生物稳态失调驱动神经炎症,肠道微生物群在AD病理生理学中的作用得到了深入的研究[28]。
3 胆固醇-胆汁酸与阿尔茨海默病
脂质通路失调与多种神经变性疾病有关,其中载脂蛋白E(APOE)ε4等位基因被确定为散发性AD最强的遗传危险因素。由于在AD患者中发现了神经元丢失和胆固醇代谢障碍,因而胆固醇及其代谢在AD发病机制中起重要作用[29-30]。通过全基因组关联研究发现胆固醇代谢相关基因(如BIN1、CLU、PICALM、ABCA7、 ABCG1及SORL1)是AD易感基因位点[31],其可能的机制是胆固醇代谢可以独立调节AD神经元中的Tau蛋白和β淀粉样蛋白,CYP46A1-胆固醇酯-Tau轴已被确定为AD早期的给药途径[32]。Varma VR等研究证实AD中胆固醇由 CYP46A1特异性酶催化的主要代谢物24S-羟基胆固醇显著降低,非酶促代谢物7α-羟基胆固醇(也可由CYP7A1酶促产生)和7β-羟基胆固醇分解代谢增加,此三种代谢物与神经炎斑块和神经原纤维病理的严重程度有关,24S-羟基胆固醇可能作为Aβ产生、tau磷酸化、神经元死亡和认知能力的调节剂发挥重要作用,胆固醇分解代谢向AD中胆汁酸合成增强的方向转变以及伴随的24S-羟基胆固醇水平降低,可能加速认知障碍进程[33]。因此阐明AD患者的胆固醇-胆汁酸代谢异常调控机制,有助于提供诊疗干预新靶点。
4 阿尔茨海默病临床和模型中的胆汁酸紊乱
病理机制一直是阿尔茨海默病研究重要的一环,有明确研究证实在AD患者的脑血管中β-淀粉样蛋白会出现沉积并造成慢性血管损伤[34],而这种损伤会导致包括BAs在内的一系列物质在大脑内的平衡发生异常。大量临床研究显示BAs浓度水平在AD患者中发生改变,其代谢紊乱可能是AD风险的潜在致病原因之一[35⇓⇓⇓-39]。
4.1 胆汁酸谱代谢紊乱与阿尔茨海默病
Mahmoudian Dehkordi S等开展一项1464例临床多中心研究[2],通过靶向和非靶向代谢组学分析20种BAs变化,结果显示AD 患者CA水平显著降低,继发性BAs及其结合形式(DCA、GDCA、TDCA和GLCA)水平升高,GDCA、GLCA和TLCA水平升高与认知功能降低显著相关,同时评估BAs水平对789名MCI患者进展为AD的风险影响,低水平CA和高水平继发性BAs比例、GDCA/CA 和 TDCA/CA 与发生 AD 痴呆的风险显著相关。有研究采用UPLC-MS/MS测定44名AD患者和94名认知正常对照受试者代谢物浓度,发现AD组多不饱和脂肪酸、酰基肉碱、色氨酸降解产物水平显著降低,BAs水平升高,在AD患者中包括CA、CDCA、UDCA、GDCA、GUDCA、别胆酸(Allocholic acid,AlloLCA)、甘氨鹅脱氧胆酸(Glycochenodeoxycholic acid,GCDCA)在内的BAs水平相较于正常对照组发生显著升高,其浓度水平与AD的严重程度呈正相关,并确定五种特异性AD标志物(CA,CDCA,AlloLCA,吲哚乳酸和色氨酸)可以区分AD与非AD[40]。Pan X等采用靶向代谢组学方法分析AD患者血浆和大脑样本中22种胆汁酸的浓度水平发现AD患者的CA含量显著降低[35]。Baloni P等使用代谢网络分析对死后脑样本分析发现AD组的初级偶联和二级BAs与CA的比例结果显示DCA、LCA、GCDCA、CDCA、AlloLCA、GDCA、TDCA和牛磺胆酸(Taurocholic acid,TCA)高于对照组,表明AD患者可能通过血脑屏障的转运受到影响[41]。Marksteiner J等针对血浆LCA水平的研究进一步揭示了AD患者体内该代谢物的水平明显高于对照组,提出LCA作为生物标志物应用于AD诊断的可能性[42]。目前已有研究提出TCA是具有细胞毒性的BAs假说,而CDCA和UDCA则被认为是一种神经保护性BAs,这种假说认为由于多种BAs含量的改变导致细胞损伤加剧及神经保护作用降低,并最终导致认知能力下降[2],因此,BAs稳态失衡与AD的发展密切相关,其代谢紊乱导致AD的具体致病机制尚在研究中。BAs谱稳态的改变可能与AD的病理生理学有关,其改变可能预示着AD发生风险增加。
Kaur H等学者在对小鼠肝脏中BAs含量分析发现,在两个不同系的AD小鼠肝脏模型中,雌性小鼠肝脏中“酸性”BAs合成途径的中间产物(3β,7α-二羟基-5-胆固醇酸)下降和CYP酶发生改变密切相关。在雄性AppNL-G-F小鼠体内偶联初级BAs,特别是牛磺酸偶联BAs(包括TDCA等)水平升高,表明AD患者体内的胆固醇和BAs合成及信号转导过程存在缺陷并对肝脏造成损伤,这项研究揭示肝脏代谢改变或BAs代谢异常可能参与AD的病理过程,肝脏中BAs代谢的变化可能是AD全身表现的一个组成部分[43]。Koike S等研究通过对AD小鼠大脑模型中BAs的测定提示CA、DCA等可作为AD诊断标志物,监测CDCA水平可对AD的症状及干预效果做出评价[44]。由此可见BAs谱的改变可能成为预示AD风险的潜在靶向生物标志物。
4.2 牛磺酸脱氧胆酸与阿尔茨海默病
牛磺酸脱氧胆酸(Tauroursodeoxycholic acid,TUDCA)是一种内源性抗凋亡BAs,由UDCA与氨基酸牛磺酸结合产生,能够透过血脑屏障,其生理作用主要是增加细胞存活及抑制神经元凋亡。目前已有研究证明TUDCA通过减少Aβ斑块累积,降低突触损失和改善认知缺陷,同时TUDCA还可消除GSK3 β过度活跃,与Tau蛋白过度磷酸化和胶质细胞激活密切相关,这种作用可能依赖于上游激酶Akt的特异性激活[45]。利用TUDCA治疗可以抑制胶质细胞的激活和增加促炎细胞的数量,降低促炎细胞因子mRNA的表达,TUDCA还被认为可减少APP/PS1小鼠模型的淀粉样低聚物累积,这是由于TUDCA增强了链脲佐菌素诱导的AD小鼠模型中的葡萄糖稳态[46]。Nunes AF等人针对其背后的机制进行了深入研究,证实TUDCA在抑制APP/PS1双转基因小鼠皮质和海马区Aβ沉积的积累及有效减少星形细胞和小胶质细胞的激活方面发挥重要作用,同时TUDCA可以增加神经元标志物MAP2、NeuN和SYN的水平[47]。因此TUDCA对AD的治疗具有良好的应用前景。
4.3 回肠胆汁酸结合蛋白与阿尔茨海默病
回肠胆汁酸结合蛋白(Ileal bile acid binding protein,I-BABP)是胆汁酸循环系统的关键组成部分,可以调节小肠胆汁酸活性,是肠上皮细胞损伤的指标[48]。Wang X等在研究中观察到MCI和AD中I-BABP水平显著升高,而I-BABP可能与T细胞协同作用并加速认知功能恶化,该研究结果提示小肠中胆汁酸的变化是导致认知功能恶化的一个促进因素,I-BABP增加与AD发病进展密切相关[49]。
4.4 其他胆汁酸代谢物与阿尔茨海默病
7α、25-二羟基-3-氧-胆酸-4-en-26-酸是一种胆汁酸前体,William J等人的研究分析了AD和痴呆患者脑脊液中的氧化甾醇和胆甾酸含量,显示AD患者组的胆汁酸前体7α、25-二羟基-3-氧-胆酸-4-en-26-酸在脑脊液中大量减少。这是由于神经元数量减少而导致的合成减少,可能与一种AD有关的干扰素刺激基因(CH25H)的表达水平有关,该基因位于10号染色体上,靠近脂肪酶A(溶酶体酸型)基因,此基因控制着与AD密切相关的水解胆固醇酯的溶酶体蛋白的表达[50],这种胆汁酸前体在调节AD的代谢方面具有关键的作用。
总结与展望
人体内BAs代谢是一个复杂的体系,其过程受到肝脏和肠道中多种酶及受体的调节,不断有证据表明BAs可能是大脑-肠道-微生物群轴内三种途径相互作用的重要调节剂,控制着包括神经、免疫和神经内分泌途径的生理功能[1,51]。BAs代谢紊乱证实与AD疾病发生和发展有着极高的关联性,而BAs与肠道微生物的相互作用同样参与其中。大量的临床实验研究支持BAs和AD病理之间具有关联性的假说,证实大脑-肠道-微生物群轴的的调控在AD病理机制研究中的意义,对 BAs信号传导和引起神经元受体的改变可能成为AD研究领域的新方向,但这些假说仍然需要进一步的临床研究进行验证,而目前关于AD各阶段中枢神经系统中BAs信号系统失调的临床研究仍然较少,需要更大样本量和多中心的实验研究为BAs水平在疾病中的调节提供数据支持,同时也是AD与BAs关联研究的重要方向。尽管如此BAs仍可被视为AD风险预防和早期诊断的生物标志物及治疗的靶点。
| [1] | Jia W, Rajani C, Kaddurah-Daouk R, et al. Expert insights: The potential role of the gut microbiome-bile acid-brain axis in the development and progression of Alzheimer's disease and hepatic encephalopathy[J]. Med Res Rev, 2020, 40(4):1496-1507. 本文引用 [3] 摘要 |
| [2] | MahmoudianDehkordi S, Arnold M, Nho K, et al. Altered bile acid profile associates with cognitive impairment in Alzheimer's disease-An emerging role for gut microbiome[J]. Alzheimers Dement, 2019, 15(1):76-92. 本文引用 [6] 摘要 |
| [3] | Baloni P, Funk CC, Yan J, et al. Metabolic Network Analysis Reveals Altered Bile Acid Synthesis and Metabolism in Alzheimer's Disease[J]. Cell Rep Med, 2020, 1(8):100138. |
| [4] | Grant SM, DeMorrow S. Bile Acid Signaling in Neurodegenerative and Neurological Disorders[J]. Int J Mol Sci, 2020, 21(17):5982. |
| [5] | Nho K, Kueider-Paisley A, MahmoudianDehkordi S, et al. Alzheimer's Disease Neuroimaging Initiative and the Alzheimer Disease Metabolomics Consortium. Altered bile acid profile in mild cognitive impairment and Alzheimer's disease: Relationship to neuroimaging and CSF biomarkers[J]. Alzheimers Dement, 2019, 15(2):232-244. 本文引用 [2] 摘要 |
| [6] | Li T, Chiang JY. Bile acid signaling in metabolic disease and drug therapy[J]. Pharmacol Rev, 2014, 66(4):948-983. 本文引用 [1] 摘要 |
| [7] | Zhou H, Hylemon PB. Bile acids are nutrient signaling hormones[J]. Steroids, 2014, 86:62-68. 本文引用 [1] 摘要 |
| [8] | Wang X, Niu Y, Yue CX, et al. Increased ileal bile acid binding protein and galectin-9 are associated with mild cognitive impairment and Alzheimer's disease[J]. J Psychiatr Res, 2019, 119:102-106. 本文引用 [1] 摘要 |
| [9] | Petrov PD, García-Mediavilla MV, Guzmán C, et al. A Network Involving Gut Microbiota, Circulating Bile Acids, and Hepatic Metabolism Genes That Protects Against Non-Alcoholic Fatty Liver Disease[J]. Mol Nutr Food Res, 2019, 63(20):e1900487. |
| [10] | Jia W, Xie G, Jia W, et al. Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis[J]. Nat Rev Gastroenterol Hepatol, 2018, 15(2):111-128. 本文引用 [1] 摘要 |
| [11] | Klingelhoefer L, Reichmann H. Pathogenesis of Parkinson disease the gut-brain axis and environmental factors[J]. Nat Rev Neurol, 2015, 11(11):625-636. 本文引用 [1] 摘要 |
| [12] | Dinan TG, Cryan JF. Gut-brain axis in 2016: Brain-gut-microbiota axis - mood, metabolism and behaviour[J]. Nat Rev Gastroenterol Hepatol, 2017, 14(2):69-70. |
| [13] | Dinan TG, Cryan JF. Gut instincts: microbiota as a key regulator of brain development, ageing and neurodegeneration[J]. J Physiol, 2017, 595(2):489-503. |
| [14] | Fiorucci S, Carino A, Baldoni M, et al. Bile Acid Signaling in Inflammatory Bowel Diseases[J]. Dig Dis Sci, 2021, 66(3):674-693. |
| [15] | Milosevic I, Vujovic A, Barac A, et al. Gut-Liver Axis, Gut Microbiota, and Its Modulation in the Management of Liver Diseases: A Review of the Literature[J]. Int J Mol Sci, 2019, 20(2):395. |
| [16] | Zhuang ZQ, Shen LL, Li WW, et al. Gut microbiota is altered in patients with Alzheimer’s disease[J]. J Alzheimers Dis, 2018, 63(4):1337-1346. |
| [17] | Vogt NM, Kerby RL, Dill-McFarland KA, et al. Gut microbiome alterations in Alzheimer's disease[J]. Sci Rep, 2017, 7(1):13537. 本文引用 [1] 摘要 |
| [18] | Jiang Y, Guo Z, Liu X, et al. Effects of Aricept on intestinal flora diversity in patients with mild Alzheimer's disease explored through high-throughput sequencing technology[J]. Am J Transl Res, 2021, 13(4):2486-2498. 本文引用 [1] 摘要 |
| [19] | Ridlon JM, Kang DJ, Hylemon PB, et al. Bile salt biotransformations by human intestinal bacteria[J]. J Lipid Res, 2006, 47(2):241-259. 本文引用 [1] 摘要 |
| [20] | Ridlon JM, Kang DJ, Hylemon PB, et al. Bile acids and the gut microbiome[J]. Curr Opin Gastroenterol, 2014, 30(3):332-338. |
| [21] | Monteiro-Cardoso VF, Corlianò M, Singaraja RR, et al. Bile Acids: A Communication Channel in the Gut-Brain Axis[J]. Neuromolecular Med, 2021, 23(1):99-117. |
| [22] | Szablewski L. Human Gut Microbiota in Health and Alzheimer's Disease[J]. J Alzheimers Dis, 2018, 62(2):549-560. 本文引用 [1] 摘要 |
| [23] | Goyal D, Ali SA, Singh RK, et al. Emerging role of gut microbiota in modulation of neuroinflammation and neurodegeneration with emphasis on Alzheimer's disease[J]. Prog Neuropsychopharmacol Biol Psychiatry, 2021, 106:110112. |
| [24] | Kesika P, Suganthy N, Sivamaruthi BS, et al. Role of gut-brain axis, gut microbial composition, and probiotic intervention in Alzheimer's disease[J]. Life Sci, 2021, 264:118627. |
| [25] | Erny D, Hrabě de Angelis AL, Jaitin D, et al. Host microbiota constantly control maturation and function of microglia in the CNS[J]. Nat Neurosci, 2015, 18(7):965-977. 本文引用 [1] 摘要 |
| [26] | Erny D, Hrabě de Angelis AL, Prinz M, et al. Communicating systems in the body: how microbiota and microglia cooperate[J]. Immunol, 2017, 150(1):7-15. |
| [27] | Kowalski K, Mulak A. Brain-Gut-Microbiota Axis in Alzheimer's Disease[J]. J Neurogastroenterol Motil, 2019, 25(1):48-60. |
| [28] | Junges VM, Closs VE, Nogueira GM, et al. Crosstalk between Gut Microbiota and Central Nervous System: A Focus on Alzheimer's Disease[J]. Curr Alzheimer Res, 2018, 15(13):1179-1190. 本文引用 [1] 摘要 |
| [29] | Feringa FM, van der Kant R. Cholesterol and Alzheimer's Disease; From Risk Genes to Pathological Effects[J]. Front Aging Neurosci, 2021, 13:690372. |
| [30] | Di Paolo G, Kim TW. Linking lipids to Alzheimer's disease: cholesterol and beyond[J]. Nat Rev Neurosci, 2011, 12(5):284-296. 本文引用 [1] 摘要 |
| [31] | Beecham GW, Hamilton K, Naj AC, et al. Genome-wide association meta-analysis of neuropathologic features of Alzheimer's disease and related dementias[J]. PLoS Genet, 2014, 10(11):e1004867. |
| [32] | Van der Kant R, Langness VF, Herrera CM, et al. Cholesterol Metabolism Is a Druggable Axis that Independently Regulates Tau and Amyloid-βin iPSC-Derived Alzheimer's Disease Neurons[J]. Cell Stem Cell, 2019, 24(3):363-375.e9. 本文引用 [1] 摘要 |
| [33] | Varma VR, Büşra Lüleci H, Oommen AM, et al. Abnormal brain cholesterol homeostasis in Alzheimer's disease-a targeted metabolomic and transcriptomic study[J]. NPJ Aging Mech Dis, 2021, 7(1):11. 本文引用 [1] 摘要 |
| [34] | Humpel C. Chronic mild cerebrovascular dysfunction as a cause for Alzheimer's disease?[J]. Exp Gerontol, 2011, 46(4):225-232. 本文引用 [1] 摘要 |
| [35] | Pan X, Elliott CT, McGuinness B, et al. Metabolomic Profiling of Bile Acids in Clinical and Experimental Samples of Alzheimer's Disease[J]. Metabolites, 2017, 7(2):28. |
| [36] | Marksteiner J, Blasko I, Kemmler G, et al. Bile acid quantification of 20 plasma metabolites identifies lithocholic acid as a putative biomarker in Alzheimer's disease[J]. Metabolomics, 2018, 14(1):1. 本文引用 [1] 摘要 |
| [37] | Mapstone M, Cheema AK, Fiandaca MS, et al. Plasma phospholipids identify antecedent memory impairment in older adults[J]. Nat Med, 2014, 20(4):415-418. 本文引用 [1] 摘要 |
| [38] | Greenberg N, Grassano A, Thambisetty M, et al. A proposed metabolic strategy for monitoring disease progression in Alzheimer's disease[J]. Electrophoresis, 2009, 30(7):1235-1239. 本文引用 [1] 摘要 |
| [39] | Olazarán J, Gil-de-Gómez L, Rodríguez-Martín A, et al. A blood-based, 7-metabolite signature for the early diagnosis of Alzheimer's disease[J]. J Alzheimers Dis, 2015, 45(4):1157-1173. 本文引用 [1] 摘要 |
| [40] | Shao Y, Ouyang Y, Li T, et al. Alteration of Metabolic Profile and Potential Biomarkers in the Plasma of Alzheimer's Disease[J]. Aging Dis, 2020, 11(6):1459-1470. |
| [41] | Baloni P, Funk CC, Yan J, et al. Metabolic Network Analysis Reveals Altered Bile Acid Synthesis and Metabolism in Alzheimer's Disease[J]. Cell Rep Med, 2020, 1(8):100138. |
| [42] | Marksteiner J, Blasko I, Kemmler G, et al. Bile acid quantification of 20 plasma metabolites identifies lithocholic acid as a putative biomarker in Alzheimer's disease[J]. Metabolomics, 2018, 14(1):1. 本文引用 [1] 摘要 |
| [43] | Kaur H, Seeger D, Golovko S, et al. Liver Bile Acid Changes in Mouse Models of Alzheimer's Disease[J]. Int J Mol Sci, 2021, 22(14):7451. |
| [44] | Koike S, Miyaji Y, Sano H, et al. Simultaneous Determination of Five Bile Acids as Potential Biomarkers for Alzheimer's Disease in Mouse Brain and Plasma[J]. Anal Sci, 2021, 37(8):1165-1170. 本文引用 [1] 摘要 |
| [45] | Dionísio PA, Amaral JD, Ribeiro MF, et al. Amyloid-βpathology is attenuated by tauroursodeoxycholic acid treatment in APP/PS1 mice after disease onset[J]. Neurobiol Aging, 2015, 36(1):228-240. 本文引用 [1] 摘要 |
| [46] | Zangerolamo L, Vettorazzi JF, Solon C, et al. The bile acid TUDCA improves glucose metabolism in streptozotocin-induced Alzheimer's disease mice model[J]. Mol Cell Endocrinol, 2021, 521:111116. |
| [47] | Nunes AF, Amaral JD, Lo AC, et al. TUDCA, a bile acid, attenuates amyloid precursor protein processing and amyloid-β deposition in APP/PS1 mice[J]. Mol Neurobiol, 2012, 45(3):440-454. 本文引用 [1] 摘要 |
| [48] | Nakahara M, Furuya N, Takagaki K, et al. Ileal bile acid-binding protein, functionally associated with the farnesoid X receptor or the ileal bile acid transporter, regulates bile acid activity in the small intestine[J]. J Biol Chem, 2005, 280(51):42283-42289. 本文引用 [1] 摘要 |
| [49] | Wang X, Niu Y, Yue CX, et al. Increased ileal bile acid binding protein and galectin-9 are associated with mild cognitive impairment and Alzheimer's disease[J]. J Psychiatr Res, 2019, 119:102-106. 本文引用 [1] 摘要 |
| [50] | Griffiths WJ, Abdel-Khalik J, Yutuc E, et al. Concentrations of bile acid precursors in cerebrospinal fluid of Alzheimer's disease patients[J]. Free Radic Biol Med, 2019, 134:42-52. |
| [51] | Mulak A. Bile Acids as Key Modulators of the Brain-Gut-Microbiota Axis in Alzheimer's Disease[J]. J Alzheimers Dis, 2021, 84(2):461-477. 本文引用 [1] 摘要 |