1 背景
脑白质高信号是在T2加权相上表现出的一种影像学信号,目前被认为可以预测中风和痴呆的风险
[1],神经病理学研究表明脑白质高信号与缺血,神经轴突丧失,神经胶质增生有关
[2-3]。目前大量观察性研究表明随着年龄增长,脑白质高信号在颅脑磁共振(MRI)上就越普遍,年龄被认为是脑白质高信号最重要的危险因素之一
[4⇓-6],因此脑白质高信号也被认为是大脑衰老的一种影像学标志。
端粒酶是染色体末端的保护DNA免受损伤的重复结构,在大多数身体组织中端粒酶长度随着时间推移而缩短
[7-8]。代表着生物学意义上的衰老
[9]。最新研究表明端粒酶缩短与中枢神经变性疾病,认知功能下降,脑缺血卒中等有密切关系
[10-11]。一些观察性研究表明白细胞端粒酶长度与WMH相关
[12-13],而也有横断面研究表明白细胞端粒酶长度与年龄相关的脑白质高信号无关
[14]。端粒酶长度所代表的生物衰老程度与脑白质高信号之间是否真的存在因果关系以及因果关系的方向在既往的研究中并没有被探讨过。
我们目前的工作采用了双向孟德尔随机化的方法,用单核苷酸多态性 (SNP) 作为工具变量,来探究端粒酶长度与脑白质高信号之间的因果关系,这种方法可以避免了观察性研究中混杂因素的干扰,以及反向因果关系的干扰。
2 材料和方法
我们选择使用两样本双向MR的方法去探究端粒酶长度和MRI T2 WMH体积之间的因果关系。正向因果关系中暴露为LTL,结局为MRI T2 WMH体积,而反向因果关系中暴露为MRI T2 WMH体积,结局为LTL。MRI T2 WMH体积的GWAS数据是来自于样本量为7705的欧洲人队列
[15]。白细胞粒酶长度的GWAS队列是来自于样本量为472174的欧洲人队列
[16]。
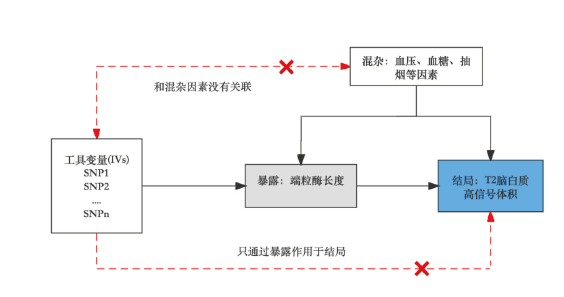
MR 应在三个基本假设下进行: (1)关联性:工具变量SNP与暴露密切相关;(2)独立性:工具变量SNP与任何潜在的混杂因素无关;(3)限制性:工具变量SNP与结果无关,除非是通过暴露的途径(图1)
[17]。
图1 孟德尔随机化的原理(以论证端粒酶长度的变化是否会导致T2脑白质高信号体积变化为例说明孟德尔研究的三大假设条件)Fig 1 Principle of Mendelian randomization (to demonstrate whether the change of telomerase length will lead to the change of T2 white matter high signal volume as an example to illustrate the three hypotheses of Mendelian study) |
Full size|PPT slide
在双向因果分析中我们选择了两组P值来筛选与暴露相关的SNP,在正向因果关系的探讨中我们使用
P = 5×10
-8为筛选暴露-LTL相关SNP的阈值。在反向因果关系的探讨中,因为工具变量数目的缺少,我们把筛选与MRI T2 WMH体积相关的SNP阈值设定为
P = 1×10
6以增加可用的工具变量SNP数目,虽然增加了MR分析中的工具变量数目,统计能力得到了提升,但是这些额外的SNP可能会违反MR分析的三个核心假设以及有存在成为弱工具变量的风险。为了得到与暴露相关独立的SNP,我们使用连锁不平衡聚类的方法(去除r>0.001和kb<10000的SNP)并选出
P值最显著的一批SNP。接着去除了不相容等位基因SNP和中等等位基因频率的回文SNP。为了剔除混杂因素的干扰,我们在phenoscanner这个网站(
http://www.phenoscanner.medschl.cam.ac.uk/)上查询了每个SNP并剔除与结局和混杂因素相关的暴露的相关SNP,从而使得我们的工具变量符合孟德尔随机化的第二条独立性和第三条限制性假设。为了避免弱工具变量的效应,我们用以下公式去计算了R2和F值。R2表示每个工具变量解释暴露的程度,MAF为次要等位基因频率,在R2中我们使用与MAF等价的EAF(效应基因频率)β值就是每个SNP对暴露的效应量,N就是暴露GWAS数据的样本量。SD是标准差,通过SE(β的标准误)计算得来。
�2=2×(1−���)×���×(���)2
��=���
接着我们使用下述的公式计算F值,其中代表每个SNP的R2值的总和,k是工具变量的数目。F值大于10 为避免弱工具变量偏倚的评估标准
[18]。
�=�−�−1�×������21−������2
我们使用了逆方差加权IVW
[19]为主要手段外加另外四种方法(分别是MR Egger, Weighted median, Simple mode, Weighted Mode)来判断暴露和结局之间的因果关系,并计算出
P值和OR值。为了保证孟德尔随机化结果的可靠性我们进行敏感性分析,我们对结果进行了异质性分析(cochran Q test)
[20] 并计算出Q-P值,该值如果大于0.05说明工具变量之间没有显著的异质性。然后我们使用MR egger-interpret的方法
[21]去检测工具变量是否有水平多效性,
P值大于0.05说明没有水平多效性存在。此外,我们使用MR-PRESSO global检验
[22]去检验水平多效性和离群值的存在。另外,我们还使用了留一法作图去判断是否单个SNP驱动主要因果关系。
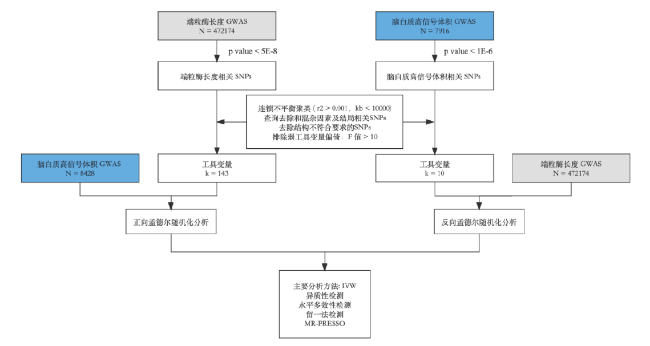
所有分析是在R4.1.4版本下进行的(
https://www.r-project.org/).MR分析涉及到的包是“TwoSampleMR”和“MRPRESSO”,如图2。
图2 验证端粒酶长度和脑白质高信号体积的双向孟德尔随机化研究流程图Fig 2 Flow chart of two-way Mendelian randomization study to verify telomerase length and high signal volume of white matter |
Full size|PPT slide
3 结果
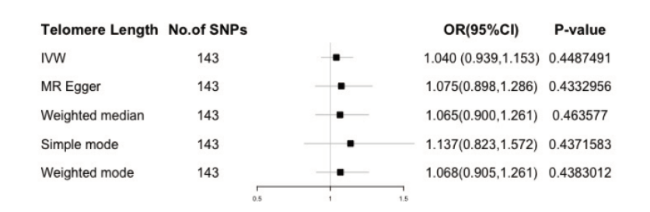
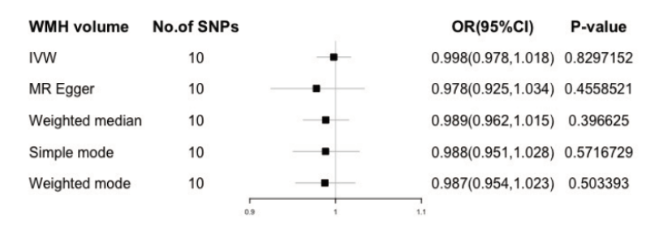
我们用双向的两样本MR研究去探究LTL和WMH之间的因果关系。以IVW为主的五种方法显示端粒酶长度和脑白质高信号之间没有因果关系(图3,4),正向和反向因果关系中选取的暴露工具变量SNP的信息在附表中。F值计算排除了弱工具变量的偏倚。Q检验显示没有异质性。根据MR-Egger test 和 MR-PRESSO global test,工具变量没有水平多效性。MR-PRESSO global test 显示没有离群值, 留一法中每个SNP的结果方向统一,没发现有单个对结果敏感的SNP,说明结果稳健可靠。
图3 端粒酶长度与脑白质高信号正向孟德尔随机化分析结果Fig 3 Results of forward Mendelian randomization analysis of telomerase length and white matter high signal |
Full size|PPT slide
图4 端粒酶长度与脑白质高信号反向孟德尔随机化分析结果Fig 4 Results of reverse Mendelian randomization analysis of telomerase length and white matter high signal |
Full size|PPT slide
4 讨论
我们使用了双向的大样本GWAS数据来验证了白细胞端粒酶长度与脑白质高信号体积之间的因果关系,我们没有发现两者之间因果关系的证据,我们的研究相比较与观察性研究,不容易受到混杂因素的干扰,而基于大样本得出的结论也避免了样本量不足带来的偏倚,Tomohisa Nezu等人的横断面研究表明端粒酶长度与年龄相关的脑白质高信号无关,而端粒酶G尾(G-tail)与脑白质高信号的严重程度相关
[14],而Kyung-Mee Choi等人的基于社区人群的横断面研究则提示了短端粒酶长度与阻塞性睡眠呼吸暂停的交互作用导致了年龄相关的脑白质高信号的发生
[23]。在另一项探究端粒酶长度和脑白质高信号的研究中发现,只有在高血压、非糖尿病和受过教育的人群中,端粒酶长度和脑白质高信号之间有显著相关性,而端粒酶长度可能是通过对于全球脑容量的保护作用从而影响老年人的认知能力
[24]。我们推断脑白质高信号并不是端粒酶长度所代表的生物学衰老这一个因素导致的,可能是在高血压、阻塞性睡眠呼吸暂停综合症共同作用下诱发的脑部小血管管壁变性导致局部脑组织缺血缺氧,未来在临床样本中需要更多相关的研究去探索衰老和其他血管危险因素在脑白质发生发展中起到的交互作用。
我们的研究有如下的局限性:1、我们只用了端粒酶长度这一个指标来探讨生物学衰老与脑白质高信号体积之间的因果关系,未来更多研究需要纳入探讨端粒酶G尾长度以及其他内源性衰老的生物学指标 2、在双样本的孟德尔随机化分析中,暴露人群和结局的人群之间可能存在样本重叠,这可能会使得结果产生偏差 3、目前研究的人群受到种族限制,均为欧洲人,结果不能推广到其他种族的人群,未来有待于在其他人种中进行验证。
总而言之,目前我们基于大样本的GWAS数据进行孟德尔随机化研究没有发现端粒酶长度和脑白质高信号的因果关系的证据,未来需要更多的研究来探究生物学衰老和脑白质高信号发生的生物学机制。


{kind=link}
{kind=link}
{kind=link}
{kind=link}