锰是人和动物体内必需的微量元素。正常情况下,锰以最适宜的水平维持机体的生理功能
[1]。近年来,由于锰的工业生产及其对环境的污染,锰的毒性越来越受到相关学者的关注
[2]。锰的过度暴露,不但会引起炎症反应、氧化应激反应,且可诱导多巴胺耗竭、神经递质释放异常、自噬功能异常、线粒体损伤、神经元的损伤等,其可能是阿尔茨海默病(Alzheimer's Disease, AD)和帕金森氏病等神经变性疾病的危险因素
[1,3-4]。锰主要聚集在星形胶质细胞和神经元细胞中,且在神经元中的毒性更强
[5]。有研究利用原代神经元培养物作为模型研究锰诱导的大脑毒性
[6]。本研究旨通过建立氯化锰(manganese chloride, MnCl
2)介导的SD大鼠大脑皮层原代神经元的氧化应激及炎症损伤模型,进一步探讨锰中毒的细胞机制及其对AD等神经变性疾病发生发展的影响。
1 材料与方法
1.1 实验动物
SPF级雌、雄SD新生大鼠,1~3 d龄,购自昆明医科大学动物中心,生产许可证:SYXK(滇)K2015-0002。
1.2 主要试剂
氯化锰(100 g/瓶)、LPS(25 mg/支)、多聚-D-赖氨酸氢溴酸盐(5 mg/支)、抗β-Actin 抗体(200 μl/支)、BSA(50 g/支)均购自美国Sigma-Aldrich公司;神经细胞培养液(500 ml/瓶)、B27(10 ml/瓶)、谷氨酰胺(100 ml/瓶)均购自美国Gibco公司;青/链霉素(100 ml/瓶)、0.25 % Trypsin-EDTA胰酶(100 ml/瓶)、蛋白酶抑制剂(1 μl/支)均购自美国Millipore公司;MAP2抗体(100 μg/支)购自美国Hyclone公司;DMEM/F-12培养基(500 ml/瓶)购自美国Hyclone公司;α-MEM培养基(500 ml/瓶)购自以色列BI公司;FBS(500 ml/瓶)购自美国Gemini公司;快速免染丙烯酰胺制胶试剂盒,10%(400 ml/套)购自美国Bio-Rad公司;抗COX-2抗体(500 μl/支)和N2(5 ml/瓶)购自美国Thermofisher公司;抗iNOS抗体(150 μl/支)购自美国BD公司;山羊抗兔IgG抗体(1 ml/支)购自美国Gene Tex公司;HRP标记的山羊抗小鼠(1 ml/支)、山羊抗兔(1 ml/支)、RIPA裂解液(100 ml/瓶)、PMSF(10 ml/支)、一氧化氮检测试剂盒(500次/盒)均购自中国碧云天公司;PBS(500 ml/瓶)购自中国Solarbio公司等。所有试剂均按照说明书配制相关工作液。
1.3 方法
1.3.1 原代神经元的培养
采用本课题组前期建立的方法
[7]。取1~3 d龄SD大鼠大脑皮层,剔除脑膜及血管,加入0.25 % Trypsin-EDTA胰酶消化10 min;过滤,用加15 % FBS和1 %青/链霉素的DMEM/F-12培养基重悬细胞,以2×10
5 cells/ml细胞密度接种至经多聚赖氨酸包被的24孔板中,置37℃、5 % CO
2培养箱中孵育4 h;细胞贴壁后,更换为加2 % B27和1 %谷氨酰胺的神经细胞培养液,每3 d换1次液,约至8 d时即可进行实验。
1.3.2 原代混合胶质细胞的培养
采用本课题组前期研究中的方法
[8]。取1~3 d龄SD大鼠大脑皮层,剔除脑膜及血管,剪碎,加入0.25 % Trypsin-EDTA胰酶消化25 min后,轻轻吹打细胞成悬液,细胞筛过滤,用含10 % FBS和1 %青/链霉素的α-MEM培养基重悬细胞后接种至T75培养瓶中,置37℃、5 % CO
2培养箱中孵育,待细胞融合时进行传代;当细胞再次融合后消化,离心,重悬,以1×10
5 cells/ml细胞密度接种至24孔板中,置37℃孵育24 h;更换为加1 % N2的无FBS同类培养基后,继续孵育约24 h后进行实验。
1.3.3 原代神经元的鉴定
采用免疫荧光染色法
[7]。将培养的原代神经元细胞悬液以1×10
4 cells/ml密度接种至经多聚赖氨酸包被的爬片上,置37℃、5 % CO
2培养箱中生长约3周时,进行鉴定:吸去上清,加入4 %多聚甲醛固定30 min;用1 % Triton-100透膜30 min;1 % BSA封闭1 h;加入MAP2抗体(1:300),置室温孵育2 h;加入山羊抗兔IgG抗体(1:1 000)避光孵育1 h;加入DAPI抗体(1:100)避光孵育20 min;封片,避光晾干,观察并拍照记录。
1.3.4 实验分组
随机将培养的原代神经元细胞分为阴性对照组(Control组)、阳性对照组(100 ng/ml LPS)及低、中、高浓度的MnCl2组(250、500、1 000 μΜ),按分组分别加入相应的药物后,置37℃、5 % CO2培养箱中孵育24 h,收集细胞培养液。以SD大鼠大脑皮层原代混合胶质细胞作为对照,观察MnCl2对原代神经元分泌NO、iNOS、COX-2的影响。
1.3.5 Griess Assay
取上述收集的细胞培养液,离心,取上清作为待测样本,按照一氧化氮检测试剂盒说明书进行操作:分别取待测样本、标准液、空白对照液各50 μl加入96孔板中,同时加入GriessⅠ和Ⅱ试剂各50 μl,置室温下孵育10 min,用酶标仪在560 nm波长下检测吸光度值,NaNO2标准曲线计算出待测样本的NO浓度。
1.3.6 Western blot
细胞孵育24 h吸去上清后,用加蛋白酶抑制剂和PMSF的RIPA裂解液提取细胞总蛋白,制备免染胶,并依次进行电泳、蛋白转至PVDF膜、1 % BSA封闭;加入抗iNOS 抗体(1:2 000)、抗COX-2 抗体(1:1 000)、抗β-Actin抗体(1:2 500),4℃孵育过夜;TBST洗膜45 min,换液2次;加入HRP标记的山羊抗小鼠抗体(1:5 000)和山羊抗兔抗体(1:5 000),置室温孵育2 h;ECL显影,Bio-Rad ChemiDoc MP成像系统显示蛋白表达水平。
1.4 统计学分析
采用Image J软件进行图像处理,用GraphPad Prism 8软件进行统计学分析,计量资料采用(均数±标准误)表示,两两比较用独立样本t 检验;组间比较用单因素方差分析(one-way ANOVA );以P < 0.05表示差异有统计学意义。
2 结果
2.1 原代神经元的生长情况
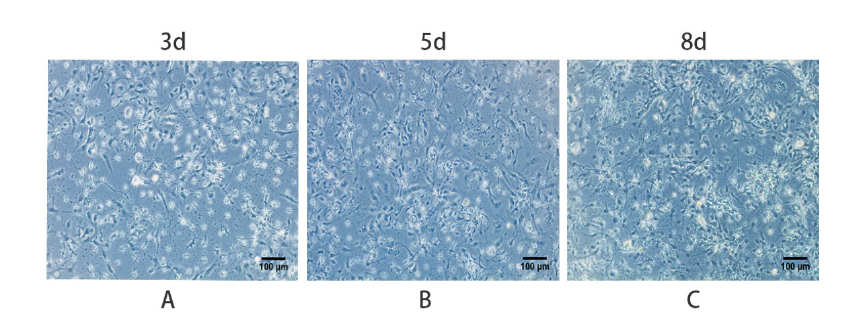
接种第一天时,现细胞漂浮,体积小且呈圆形透明状,无突起;细胞培养3 d后,胞体呈类圆形,部分细胞有细小突起;细胞培养5 d后,胞体进一步长大,细胞突起增多开始相互连接呈稀疏网状。细胞培养8 d后,神经元胞体生长饱满,细胞突起较多,形成网络状分布,见图1。
图1 原代神经元的生长情况Fig. 1 Growth of primary neurons |
Full size|PPT slide
2.2 原代神经元鉴定结果
红色荧光标记的MAP2与神经元胞体和树突结合后呈红色(图2A);胞核与DAPI抗体结合后呈蓝色,为细胞总数(图2B);二者融合后,呈1个完整的神经元(图2C箭头所示)。以Merge/DAPI×100 %,计算出神经元阳性率为82.63 %。
图2 免疫荧光染色法(MAP2抗体)鉴定培养的原代神经元Fig. 2 Immunofluorescence staining of primary neurons by MAP2 antibody |
Full size|PPT slide
2.3 MnCl2对原代神经元分泌NO浓度的影响
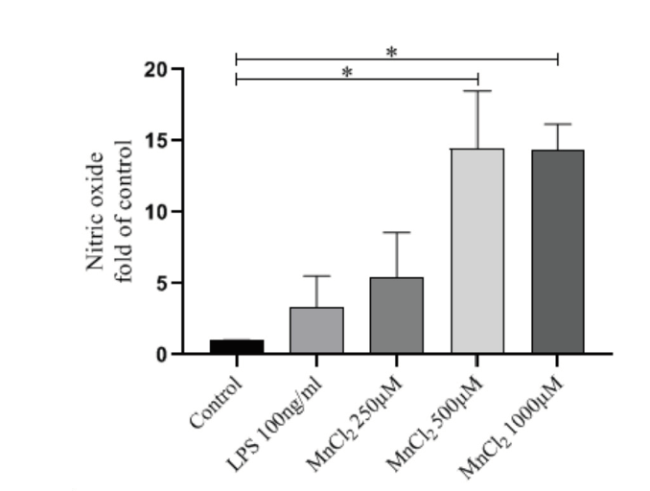
结果显示,不同浓度的MnCl2和100 ng/ml的LPS与原代神经元共孵育24 h后,NO浓度均高于Control组,且MnCl2组NO浓度明显高于LPS组。当原代神经元暴露在500或1 000 μM的MnCl2时,NO浓度明显高于Control组,差异均有统计学意义(P < 0.05, n=3),见图3。
图3 不同浓度的MnCl2对原代神经元分泌NO浓度的影响Fig.3 Effect of different concentrations of MnCl2 on NO produced by primary neurons Note: * P < 0.05, n=3 |
Full size|PPT slide
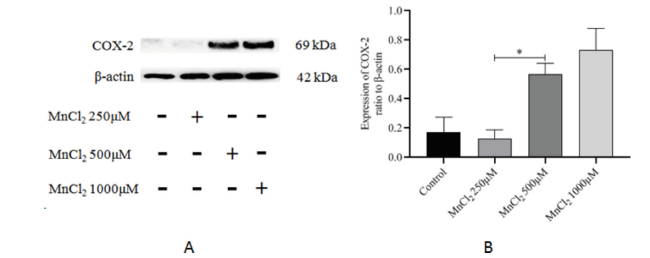
2.4 MnCl2对原代神经元iNOS及COX-2蛋白表达水平的影响
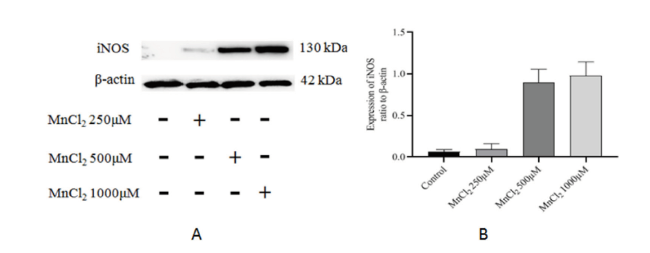
结果显示,不同浓度的MnCl2与原代神经元共孵育24 h后,iNOS及COX-2蛋白表达水平随MnCl2浓度的升高而增加,但与Control组比较,差异均无统计学意义(P均 > 0.05, n= 3),见图4、图5。
图4 不同浓度的MnCl2对原代神经元iNOS蛋白表达的影响Fig. 4 Effect of different concentrations of MnCl2 on the protein expression of iNOS by primary neurons |
Full size|PPT slide
图5 不同浓度的MnCl2对原代神经元COX-2蛋白表达的影响Fig. 5 Effect of different concentrations of MnCl2 on the protein expression of COX-2 by primary neurons Note: * P < 0.05, n=3 |
Full size|PPT slide
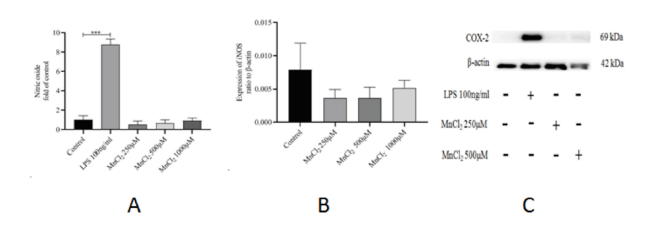
2.5 MnCl2对原代混合胶质细胞NO、iNOS和COX-2表达的影响
原代混合胶质细胞分别与250、500、或1 000 µM的MnCl2共孵育24 h后,NO、iNOS表达水平无明显变化(P > 0.05, n=3)(图6A、B);250或500 μM的MnCl2与原代混合胶质细胞共孵育24 h后,COX-2蛋白表达也无明显变化(图6C)。100 ng/ml的LPS与原代混合胶质细胞共孵育24 h后,NO明显上调,差异有统计学意义(P < 0.000 5, n=3)(图6A);COX-2蛋白表达也明显上调(图6C)。表明原代混合胶质细胞仅对LPS产生反应,而对不同浓度的MnCl2不产生反应。
图6 不同浓度的MnCl2对原代混合胶质细胞NO、iNOS和COX-2蛋白表达的影响Fig. 6 Effects of different concentrations of MnCl2 on the production of NO, iNOS, COX-2 by primary mixed glia Note: ***P < 0.000 5, n =3 |
Full size|PPT slide
3 讨论
锰对机体内的平衡调节可以被视为生物锰状态和最佳大脑健康的倒“U”形关系
[1]。正常人脑锰浓度为20.0~52.8 μM
[9],从豆类、坚果、大米、绿色蔬菜等含锰食物中摄取的锰足够满足人和动物的生理需求,故锰缺乏并不常见。但由于我国是锰铁合金的主要生产国,长期过量暴露于锰职业的人员数量庞大,加之含锰汽油抗爆剂、含锰杀虫剂等的大量使用,导致饮水和食物的锰污染,所以普通人群可能存在超过生理需求的锰暴露,因此锰暴露越来越受到相关学者的关注
[2]。 相关的研究结果显示,锰不但能诱导多巴胺能神经元产生氧化应激和炎症神经毒性反应
[12];慢性锰暴露还可导致锰在小鼠大脑皮层中积累,诱导皮质神经元功能障碍,影响工作记忆
[13],还会加剧大鼠小脑皮层的神经元改变、氧化/亚硝化应激和促炎细胞因子释放,降低抗氧化活性
[14];锰代谢紊乱可导致神经系统损伤
[1,10,11]。
NO是一种信号分子,参与机体很多重要生理过程,而内源性的NO是由一氧化氮合酶(nitric oxide synthase, NOS)催化氧化L-精氨酸产生。NOS主要分为诱导型iNOS、神经元型nNOS和内皮型eNOS 3种,其组织分布和生物学作用各不同,其中iNOS在机体被刺激后表达并催化合成非生理浓度的大量的NO
[16]。当NO释放过多时,即可引起神经元产生病理变化,产生神经毒性
[17]。Kaur 等
[18]研究发现,一氧化氮参与锰诱导Wistar大鼠产生神经毒性。
COX-2是一种重要的炎症介质,主要分布于中枢神经系统的大脑皮质神经元、海马神经元和其他端脑的树突及树突棘中,其与许多中枢神经系统疾病的病理过程中神经元的变性有关
[19]。Li等
[20]以15 mg/(kg·d)剂量的MnCl
2腹腔注射SD大鼠,每周5 d,连续12周,结果发现大鼠大脑皮层中COX-2等炎症因子的mRNA和蛋白表达水平均显著增加;Nkpaa 等
[14]研究也发现,大鼠在30 mg/kg锰暴露35 d后,其小脑皮层中的NO水平和COX-2蛋白表达水平明显增加。神经元为锰中毒最先累及的对象,易受锰诱导产生氧化应激产物和炎症因子,如iNOS、NO、COX-2等,导致神经元的丢失,进而诱发AD
[1,3,21],增加AD的患病风险
[5,19]。
在前期的研究中,本课题组已成功培养了SD大鼠大脑皮层原代神经元,并用LPS和低聚Aβ
1-42介导SD大鼠大脑皮层原代神经元NO和COX-2指标的上调,成功建立了氧化应激及神经炎症反应模型
[7],为本研究奠定了基础。本研究根据前期原代神经元的培养方法,改用MnCl
2介导SD大鼠大脑皮层原代神经元产生氧化应激和炎症反应,建立了神经元损伤模型,采用免疫荧光染色对原代神经元进行鉴定,计算得出MAP2标记阳性率为82.63 %。
本研究结果与上述相关研究结果相似
[14,18,20],即不同浓度的MnCl
2均可介导SD大鼠大脑皮层原代神经元NO浓度上调,且当MnCl
2浓度达到500或1 000 μM时,NO浓度明显高于Control组,也明显高于阳性对照组(100 ng/ml LPS),差异均有统计学意义(
P < 0.05),表明锰对SD大鼠大脑皮层原代神经元具有较大的毒性作用,其有可能是AD和帕金森氏病等神经变性疾病的危险因素;MnCl
2介导的SD大鼠大脑皮层原代神经元iNOS和COX-2蛋白表达水平虽然随着MnCl
2浓度的增加而升高,但与Control组比较差异无统计学意义(
P > 0.05)。
本研究还用不同浓度的MnCl2介导了SD大鼠大脑皮层原代混合胶质细胞,结果显示,NO及iNOS、COX-2表达水平均无明显变化,表示MnCl2对SD大鼠大脑皮层原代混合胶质细胞的NO及iNOS、COX-2无介导作用。
本研究成功地建立了MnCl2介导的SD大鼠大脑皮层原代神经元的氧化应激及炎症反应模型,即原代神经元锰暴露的实验室模型,为探索防治锰中毒的药物研究提供了一种新的细胞模型、理论基础及细胞分子生物学依据。本研究结果也提示,在日常生活中,存在锰暴露风险的人群,在做好自我防护的同时,注重大脑健康,避免大脑神经元因锰中毒带来的损伤,避免AD等神经变性疾病的发生。
由于时间短等原因,本研究暂未进行神经元锰易感性机理的研究,为此,我们将在后续的实验中进行进一步的深入研究。



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}