
2022年第一期

6-OHDA诱导帕金森病模型小鼠多巴胺能神经元坏死性凋亡的可能机制
日期:2023-11-22
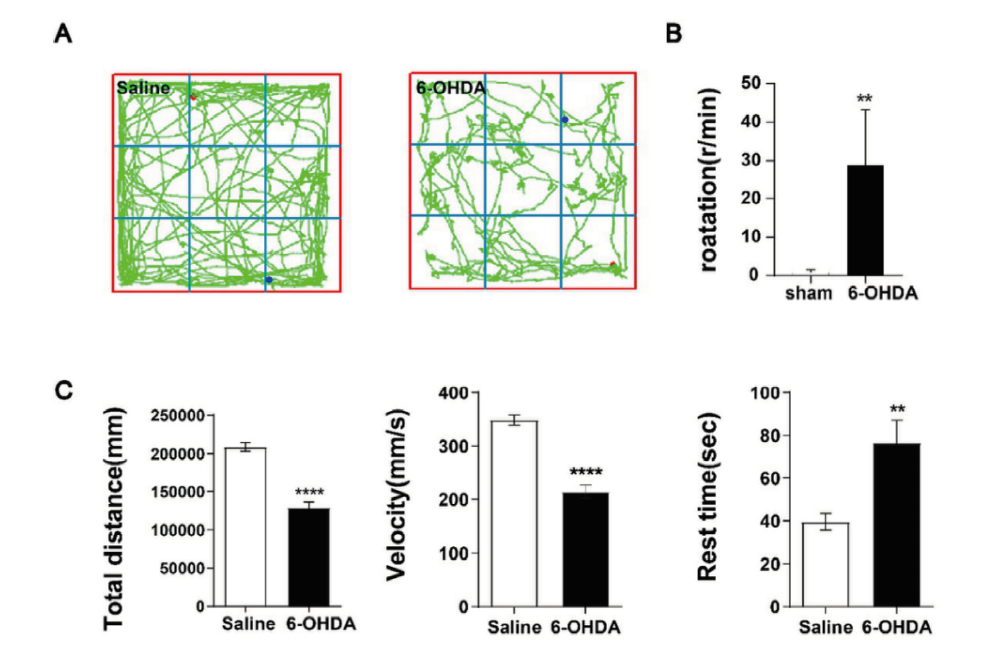
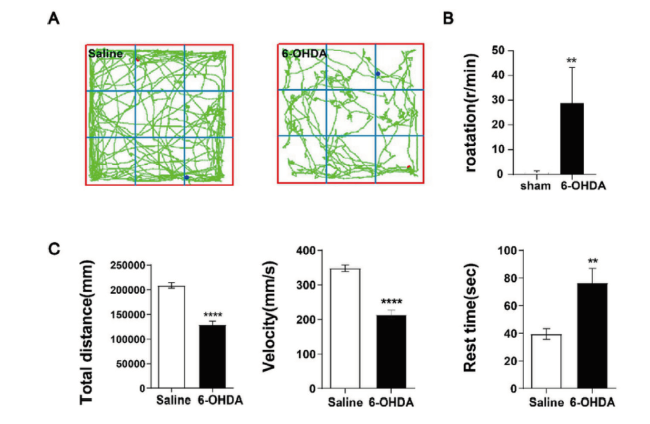
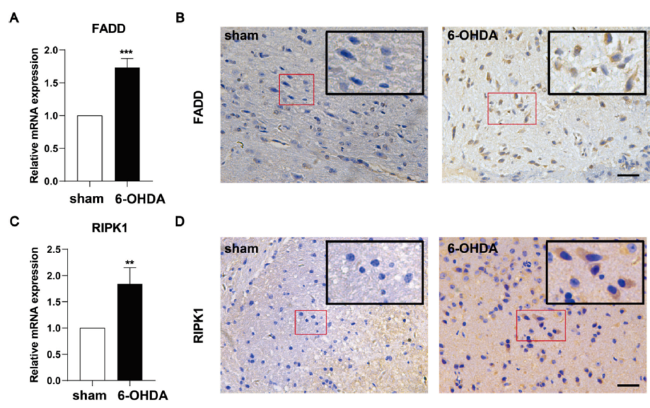
目的: 探讨6-羟基多巴胺氢溴酸盐(6-hydroxydopamine,6-OHDA)诱导帕金森病(parkinson's disease,PD)模型小鼠多巴胺能神经元坏死性凋亡的细胞机制。方法: 将含0.02% 抗坏血酸和0.9% 生理盐水的6-OHDA注入开颅后的8周龄、健康雄性C57/BL小鼠的单侧黑质(substantia nigra,SN) 区域构建PD小鼠模型(模型组),用APO旋尾实验和旷场实验验证PD小鼠模型的构建情况;取模型组小鼠脑组织进行免疫组化(immunohistochemistry,IHC)检测多巴胺能神经元(dopaminergic neurones,DA)标记物酪氨酸羟化酶(tyrosine hydroxylase, TH)恢复水平;通过qPCR和IHC检测模型组小鼠坏死性凋亡相关基因(FADD,RIPK1,TRADD)mRNA和蛋白表达水平。以左脑黑质区域只注入含0.02%抗坏血酸的0.9%生理盐水的小鼠作为正常对照组(假手术组)。结果: 与对照组比较,6-OHDA组小鼠的旋转次数明显增加,跑动距离和平均速度明显减少(P < 0.05),静止时间则明显增加(P < 0.05);6-OHDA组小鼠纹状体区域(STR)和黑质区域(SN)的TH阳性表达均明显降低(P < 0.01),其中STR中的致密神经树突纤维明显减少,SN中的DA神经元大量丢失,仅可见少量多巴胺能神经元细胞存留,残留的阳性神经元细胞胞体小,突起短;6-OHDA组小鼠RIPK1、FADD的基因和蛋白表达水平均明显增加。结论: 6-OHDA可促进DA能神经元变性死亡,其可能机制是通过促进FADD-RIPK1坏死性凋亡信号通路。
Objective: To investigate the cellular mechanism of 6-hydroxydopamine hydrobromide (6-OHDA) induced necrotic apoptosis of dopaminergic neurons in a parkinson's disease (PD) model mouse.Method: The model of Parkinson's disease was constructed by injecting 6-OHDA containing 0.02% ascorbic acid and 0.9% saline into the unilateral substantia nigra (SN) region of 8-week-old, healthy male C57/BL mouse after craniotomy, and the construction of the PD mouse model was verified by the APO spin tail test and the open field test. Brain tissues from model mouse were subjected to immunohistochemistry (IHC) to detect the recovery of dopaminergic neurones (DA) marker tyrosine hydroxylase (TH); the expression levels of FADD, RIPK1 and TRADD were detected by qPCR and IHC. The mRNA and protein expression levels of necrotizing apoptosis-related genes (FADD,RIPK1,TRADD) were measured by qPCR and IHC. A group of mouse injected with only 0.9% saline containing 0.02% ascorbic acid in the nigrostriatal region of the left brain was used as the normal control group. Result: Compared with the control group, mouse in the 6-OHDA group showed a significant increase in the number of rotations, a significant decrease in running distance and mean speed (P < 0.05), and a significant increase in resting time (P < 0.05). The positive expression of TH was significantly reduced in both striatal region (STR) and substantia nigra (SN) of mouse in the 6-OHDA group (P < 0.01), where dense nerve dendritic fibers were significantly reduced in STR and a large number of DA neurons were lost in SN, and only a few dopaminergic neuronal cells were seen to survive, with small cytosomes and short protrusions in the remaining positive neuronal cells. The gene and protein expression levels of RIPK1 and FADD were significantly increased in the 6-OHDA group of mouse. Conclusion: 6-OHDA promotes the degenerative death of Dopaminergic neurons by a possible mechanism that promotes the FADD-RIPK1 necroptosis signaling pathway.
帕金森病 / 坏死性凋亡 / 6-羟基多巴胺氢溴酸盐 / FADD / RIPK1
Parkinson's disease / Necrotic apoptosis / 6-OHDA / FADD / RIPK1

{kind=link}
{kind=link}
{kind=link}