
2023年第三期

载脂蛋白E调控阿尔茨海默病病理的机制
日期:2023-11-23
阿尔茨海默病(Alzheimer's disease,AD)的病理特征是淀粉样斑块(amyloid plaques)和神经原纤维缠结(neurofibrillary tangles,NFTs)的沉积。而淀粉样斑块的重要成分是β淀粉样蛋白(Aβ)。载脂蛋白E基因(APOE)是AD最主要的风险基因之一,其编码的载脂蛋白E(APOE),被认为与AD的多种致病过程有关。人类APOE有三种主要的等位基因:APOEɛ2、APOEɛ3和APOEɛ4。其中APOEɛ4可以促进小胶质细胞和星形胶质细胞的炎症反应和相关细胞因子的分泌,继而引发神经炎症。APOE还可直接与Aβ相互作用,促进其在不溶性纤维沉积物中的聚集和沉积。除了参与Aβ病理,来自星形胶质细胞和神经元的APOE4通过不同的方式,促进了tau蛋白的高度磷酸化以及病理性tau蛋白介导的神经元死亡和神经变性病变。此外,APOE4伴随着更多的补体激活导致的突触丢失。神经毒性APOE4片段可引起的线粒体功能障碍和血脑屏障(blood brain barrier, BBB)通透性的增加。髓样细胞-2上表达触发受体(Triggering receptor expressed on myeloid cell 2, TREM2)依赖途径上调的APOE转录,可诱导小胶质细胞参与的神经炎症反应。微生物菌群对神经炎症的调节也通过APOE依赖的方式。总之,APOE对AD的发生及进展产生重要的影响,以APOE/APOE受体或其聚合物以及其保护性突变等为靶点的治疗,可能为AD的治疗提供新的思路。
Abstract
Alzheimer’s disease (AD) is pathologically characterized by the accumulation of amyloid plaques and the deposition of neurofibrillary tangles (NFTs). An important component of amyloid plaques is β amyloid protein (Aβ). Apolipoprotein E genotype (APOE) has been reported as the biggest genetic risk factor for AD and apolipoprotein E (APOE), the APOE gene-encoded protein, has been suggested to be involved in a variety of pathogenic processes of AD. Human APOE has three major allelic variants: APOE ɛ2, APOE ɛ3, and APOE ɛ4. APOE alleles, especially APOE ɛ4, promoted inflammatory response and secretion of related cytokines in microglia and astrocytes. APOE interacts with Aβ and promotes its aggregation and deposition in insoluble fibrillar deposits. Beside the Aβ pathology, APOE4 from astrocytes and neurons could facilitate tau phosphorylation, as well as tau-mediated neurodegeneration. In addition, APOE4 is accompanied by greater complement activation and subsequently loss of synapse. Mitochondrial alterations and an increase of blood-brain barrier (BBB) permeability could be caused by neurotoxic APOE4 fragments. In AD, a triggering receptor expressed on myeloid cell 2 (TREM2)-dependent pathway could activate APOE transcription, which induces the activation of microglia. The regulation of neuroinflammation by microbial communities is also dependent on APOE. In conclusion, APOE has an important impact on the mechanisms of AD, and therapies targeting APOE/APOE receptors or polymers, as well as protective variants, may provide new ideas for the treatment of AD.
关键词
载脂蛋白E / 阿尔茨海默病 / 病理机制 / β淀粉样蛋白 / Tau蛋白
Key words
Apolipoprotein E / Alzheimer’s disease / Pathological mechanism / Aβ / Tau
引用本文
王晓彤 , 王超 , 赵颖俊. 载脂蛋白E调控阿尔茨海默病病理的机制. 阿尔茨海默病及相关病杂志. 2023, 6(3): 241-248 https://doi.org/10.3969/j.issn.2096-5516.2023.03.009
Xiaotong WANG , Chao WANG , Yingjun ZHAO. Apolipoprotein E regulates the pathology of Alzheimer's disease. Chinese Journal of Alzheimer's Disease and Related Disorders. 2023, 6(3): 241-248 https://doi.org/10.3969/j.issn.2096-5516.2023.03.009
1 APOE概述
阿尔茨海默病(Alzheimer's Disease, AD)是最常见的痴呆类型,通常为散发性的,并常见于65岁以上的老年人,因此也被称为迟发性AD[1]。AD在临床上多表现为进行性的记忆力减退和认知功能退化等[2],其神经病理学特征主要表现为患者大脑中具有淀粉样斑块(amyloid plaques)沉积形成的胞外淀粉样斑块沉积和高度磷酸化的tau蛋白于胞内沉积形成的神经原纤维缠结(neurofibrillary tangles,NFTs)。而淀粉样斑块的重要成分就是β淀粉样蛋白,即Aβ。AD是一种具有高度遗传性的神经变性疾病,全基因组关联性分析(Genome wide association study, GWAS)和meta分析鉴定出了30多个AD相关的风险基因[3⇓-5]。其中载脂蛋白E基因(APOE)是与迟发性AD关联性最高的风险基因之一,其含有三种主要的等位基因:APOEɛ2、APOEɛ3和APOEɛ4[6,7]。研究表明,APOEɛ4(与最常见的APOEɛ3等位基因相比)是目前发现的迟发性AD最强的遗传风险因素,而相对罕见的APOEɛ2等位基因是迟发性AD的遗传保护因素[4,8]。具体来说,有两个APOEɛ4拷贝的人患AD的风险比普通人高出12~20倍[9]。而APOEɛ2等位基因具有一定的保护作用,APOE2的携带者约为7%[10]。APOEɛ4携带者的Aβ斑块负荷较高,脑淀粉样血管病变更严重,而APOEɛ2携带者的Aβ斑块负荷较低。除AD外,APOEɛ4还与非痴呆个体年龄相关认知能力下降的风险以及其他神经变性疾病有关,如路易体痴呆和额颞叶痴呆等[11⇓⇓⇓-15]。
APOE基因编码的载脂蛋白E(Apolipoprotein E,APOE),是低密度脂蛋白受体(Low-density lipoprotein receptor, LDLR)家族的配体[16]。在生理条件下,大脑中的APOE主要由星形胶质细胞合成;在神经炎症等病理情况下,小胶质细胞和神经元也会参与APOE的合成。通过与神经细胞表面的受体结合,APOE承担着将星形胶质细胞合成的胆固醇和磷脂运输至神经元的任务,维持着大脑中的脂质代谢稳态。同时,APOE还可以参与多种免疫调节和炎症反应等。作为APOE在大脑中最主要的细胞表面受体,LDLR家族成员在各个神经细胞中都有表达,具有较短的细胞内结构域、跨膜结构域和较大的胞外结构域。不同的LDLR家族成员具有不同的生理作用,这可能是由于它们与配体的亲和力、信号效力、细胞定位、表达模式和内吞率不同导致的[17]。APOE的三种等位基因编码的不同APOE蛋白亚型仅仅具有1~2个氨基酸的差异:APOE2(半胱氨酸[Cys]112,Cys158)、APOE3(Cys112,精氨酸[Arg]158)和APOE4(Arg112,Arg158)[18],亚型之间的氨基酸差异会影响脂质结合和受体结合[19]。除此之外,APOE还存在很多变体,APOE3 R145C 错义变体与AD风险增加有关[20],而另一种罕见变体,APOE V236E (APOE3-Jacksonville),与保护作用有关[21]。近期发现的研究还发现携带两个APOE3 R136S(APOE3-Christchurch)突变的人的认知功能可以得到一定程度的保护[22]。大量的研究表明,APOE可以通过多个途径和多种细胞调控AD病理的发生发展。APOE是一种Aβ结合蛋白[23⇓⇓⇓-27],以亚型依赖的方式调节Aβ聚集和清除途径[28-29]。APOE还被认为可调节tau蛋白的过度磷酸化和扩散,并且APOE的水平与认知障碍患者脑脊液(Cerebrospinal fluid, CSF)中的tau蛋白水平呈正相关[30]。此外,APOE在神经炎症和后续途径中起着重要作用[31],例如神经毒性和线粒体功能障碍等[32-33]。此外,中枢神经和外周系统中的APOE可能通过破坏两侧血脑屏障(blood-brain barrier, BBB)的完整性而影响AD的疾病进程[34]。
2 APOE参与神经炎症调控AD的发病机制
小胶质细胞是神经炎症的最大参与者,而APOE是胶质细胞炎症反应的主要调节因子之一,能够多途径的影响胶质细胞的功能和AD期间反应性胶质细胞的积聚(胶质增生)等[35⇓-37]。有研究发现,携带APOEɛ4的5xFAD淀粉样变性小鼠深皮质层小胶质细胞过度增生,并且伴有更多营养不良小胶质细胞出现[38]。此外,当不同亚型的人源APOE敲入小鼠与Tau模型(P301S)鼠杂交时,携带APOEɛ4的小鼠海马和内嗅/梨状皮质中的CD68阳性小胶质细胞负荷明显增加[39],说明APOEɛ4参与了tau病理的异常神经胶质反应。为了模仿AD中的炎症反应,研究人员使用脂多糖(Lipopolysaccharide, LPS)刺激小胶质细胞,经过APOE4和LPS处理过的小胶质细胞能释放大量促炎因子,如肿瘤坏死因子-α(Tumor necrosis factor-α,TNF-α)和白细胞介素等以及其他可能导致周围组织损伤的因素[40]。而APOE敲除小鼠的小胶质细胞则分泌较低水平的抗炎因子[41],表明APOE亦可促进小胶质细胞抗炎因子的分泌,从而在中枢神经系统稳态中发挥生理作用。但是中枢神经系统稳态在AD中被破坏,APOEɛ4的小胶质细胞的促炎作用强于抗炎作用[42]。体外实验中,多能干细胞诱导分化的APOEε4小胶质细胞也表现出促炎基因高表达和对Aβ的吞噬功能受损[39],一旦炎症反应失控,促炎因子的增加将会损害周围的神经元,导致神经元变性。
除了小胶质细胞,星形胶质细胞也引起神经炎症[43]。在携带APOEε4的P301S Tau转基因小鼠中,星形胶质细胞的促炎基因显著上调[39],表明APOE ε4对星形胶质细胞的神经炎症产生重要影响。多能干细胞诱导分化的APOEε4星形胶质细胞,表现出了胆固醇代谢紊乱和对Aβ吞噬能力减弱[44],神经营养支持减少[45],突触修剪受损[46]等一系列重要生理功能的紊乱。相比于APOEɛ4星形胶质细胞,APOEɛ4星形胶质细胞的溶酶体失调导致Ca2+兴奋性增加,膜脂质体和细胞内胆固醇分布改变[47]。经LPS处理之后,APOEɛ4星形胶质细胞能产生更多趋化因子配体3(C-C Motif Chemokine Ligand 3,CCL-3)[48],进而引起小胶细胞神经炎性级联反应[41]。小胶质细胞和星形胶质细胞相互作用,共同增强AD中神经炎性反应[31]。
以上研究说明,小胶质细胞和星形胶质细胞之间存在相互作用,一旦小胶质细胞被激活,它们就可以释放细胞因子,进一步促进星形胶质细胞的激活,从而形成一个对周围环境有害的前馈回路。而星形胶质细胞也可因趋化因子表达上调,模拟出促进小胶细胞神经炎性反应的条件。因此,小胶质细胞和星形胶质细胞之间的紧密联系共同增加了AD中的神经炎性反应。
3 APOE调控AD主要病理的机制
3.1 APOE调控Aβ病理的机制
脑内的Aβ积聚是AD发病主要病理特征,而APOE/APOE受体在维持Aβ的聚集和清除中发挥着重要作用。早期研究发现致密的Aβ斑块中含有APOE[49],APOE亚型之间的构象差异导致其与Aβ的亲和力不同,其中APOE4和Aβ的亲和力最高。APOEɛ4基因敲入小鼠也表现出更高的Aβ斑块负荷[50],而用针对APOE4的抗体,靶向斑块中APOE4的免疫疗法可减少小鼠大脑Aβ的沉积[51]。选择性的抑制星形胶质细胞中的APOE4会导致Aβ斑块沉积大幅减少,斑块致密程度下降,同时,淀粉样斑块周围疾病相关小胶质细胞标志物(C-type lectin domain family 7 member A,Clec7a)和APOE的表达较低,表明小胶质细胞活化减少[52]。在研究人脑组织样本时,研究者发现APOE4还能促进可溶性Aβ寡聚体(oligomeric Aβ,oAβ)增加,而毒性相对较高的oAβ与突触结合会加剧斑块周围的突触丢失[53]。低密度脂蛋白受体蛋白1(Low density lipoprotein receptor-related protein 1 ,LRP1)介导是清除Aβ的主要途经,而APOE可与Aβ竞争性结合LPR1[29],从而影响神经元、胶质细胞和内皮细胞等细胞对Aβ的清除[54-55]。不仅如此, APOE4还能显著促进Aβ肽进入积聚状态[56-57],抑制Aβ的清除和Aβ降解酶的降解作用[58]。体外研究显示,APOE4的存在使得小胶质细胞对凋亡神经元的吞噬作用增强,但同时对Aβ的清除率反而降低,这种特异性增强吞噬凋亡神经元的作用促进了小胶质细胞从静息态转变为有害的活化表型[59]。除此之外,APOE近期被发现可与ɤ-分泌酶相互作用,并通过其保守的C末端区域以底物特异性的方式抑制其在细胞内的作用。这种载脂蛋白C末端介导的抑制活性在不同的载脂蛋白异构体中被不同程度地削弱,具体表现为抑制程度APOE2 >APOE3 > APOE4,与他们相关的AD风险成反比。于此同时,在AD小鼠模型中,神经元中APOE的C末端区域会迁移到位于海马下托(subiculum)的淀粉样蛋白斑块区域并减轻斑块负担。总而言之,载脂蛋白E的这种精准的ɤ-分泌酶抑制作用可能会降低散发性AD的风险[60]。
3.2 APOE调控tau病理的机制
除了参与Aβ依赖性神经变性以外,APOE还可以通过调控神经炎症反应,进而影响AD疾病进程中tau蛋白病理的发生和发展,在携带不同APOE亚型的P301S Tau转基因小鼠中,由病理性tau蛋白介导的神经变性病变呈现出APOE亚型依懒性(APOE4 >APOE3 > APOE2)。于此同时,APOE的缺失则极大程度地降低了tau蛋白病病理,暗示了在tau蛋白病病理进程中,APOE可能部分通过毒性增益的方式参与。特异性地敲除星形胶质细胞表达的APOE4,而不是APOE3,可以显著降低由病理性tau蛋白介导的神经炎症及神经变性病变[61]。此外,高度磷酸化的微管相关蛋白tau对神经元具有毒性作用,是神经原纤维缠结的主要成分,而APOE已被证明是这种tau介导的神经元死亡发生的必要条件[39]。在转基因小鼠模型中,APOE4在受损神经元中过度表达,促进了tau磷酸化以及tau介导的神经元死亡[62]。而从神经元中选择性敲除APOEε4可以显著降低tau病理、胶质增生、神经变性变、神经元过度兴奋和髓鞘缺失[63]。同时,单核RNA测序显示,去除神经元APOE4后,大大减少了神经变性疾病相关的神经元亚群、 少突胶质细胞、星形胶质细胞和小胶质细胞,从而显示了它们的积累与tau病变的严重程度相关联[63]。APOE4还被证明通过体内的tau依赖机制损害GABA能中间神经元,从而导致其功能障碍和最终死亡[64-65]。
4 APOE通过其他途径调控AD发病的机制
目前认为AD早期的突触丢失部分是由于异常的小胶质细胞和星形胶质细胞补体介导的吞噬作用[66-67]。APOE4伴随着更多的补体激活[68],异常的胶质细胞中补体介导的吞噬作用会导致突触丢失。此外,受损的线粒体可能在AD的发病机制中发挥基本作用[69]。在培养的神经细胞系中发现,APOE C末端截短片段的受体结合区使其逃避分泌,脂质结合区介导线粒体的功能障碍[70];对人脑组织分析发现,APOEɛ4携带者的细胞色素氧化酶活性降低,也提示线粒体功能障碍[71]。近期研究人员发现,与没有携带APOEɛ4的AD患者相比,携带APOEɛ4的AD患者在血液中显示出较高的羟基自由基水平和较低的脑氧利用率[72-73]。氧化应激水平的增加,可能会导致AD的提前发生[74]。
另一个日益受到关注的领域是APOEε4等位基因对血脑屏障的影响,APOEɛ4携带者中Aβ异常成像的可能性是正常人的两倍,这种异常成像是由抗体介导的Aβ从脑实质进入血流引起的,提示血脑屏障通透性增加[75]。APOE4还可以通过非Aβ依赖的方式增加血脑屏障通透性[76]。神经病理学研究还表明,Aβ斑块清除与Aβ和APOE从斑块到血管的重新分布以及更严重的脑淀粉样血管病变相关的血管病变有关[77]。周细胞是APOE的另一个细胞来源,正因其在脑淀粉样血管病发病机制中的意义而受到关注。在Aβ沉积的小鼠模型中,周细胞丢失导致脑淀粉样血管病和Aβ斑块增加[78]。周细胞通过LRP1以APOE亚型依赖的方式吸收Aβ,与E3相比,APOE4干扰了这种摄取[55]。人APOEε4周细胞表达的APOE mRNA和蛋白质水平高于APOEε3的周细胞,导致Aβ血管积聚增加[79]。
APOE和髓性细胞-2上表达的触发受体(triggering receptor expressed on myeloid cell 2, TREM2)均参与小胶质细胞对斑块的趋化作用[80],TREM2在中枢神经系统中由小胶质细胞大量表达,并参与小胶质细胞的关键功能,包括吞噬、细胞因子释放、脂质感应、小胶质细胞增殖和迁移[81⇓⇓⇓-85]。在AD早期,APOE的上调不依赖于TREM2,但随后一条TREM2依赖的途径激活了APOE转录,从而诱导小胶质细胞引起的炎症和变性加剧[86]。新的证据表明,肠道微生物群以一种APOE亚型依赖的方式调节神经炎症。研究人员用了表达人类APOE异构体的tauopathy基因模型小鼠,用抗生素扰乱其肠道微生物菌群可以一定程度上逆转胶质增生、tau病理的进展,这种情况在APOEε3的tau病理小鼠身上更为明显,相比之下,在APOEε4的tau病理小鼠中,tau介导的神经变性变并没有因为抗生素的治疗而发生很大的改变,可能是APOE4的毒性和对大脑局部先天性免疫反应的影响比APOE3的强,最终没能使神经变性病变的保护作用达到相同的程度[87]。针对肠道微生物菌群的治疗,可以为遏制AD及tau病理的进展提供新的思路[88-89] 。见图1。
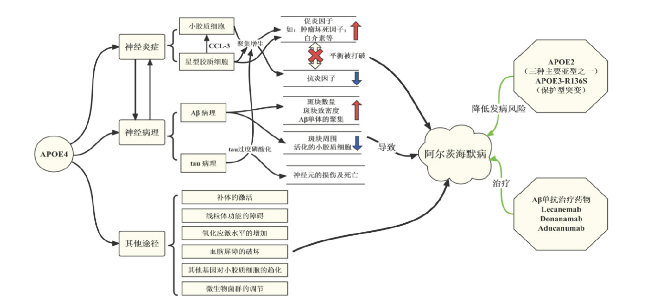
{kind=link}
5 总结与展望
APOE是AD的风险基因,由APOE基因编码的APOE蛋白在AD的发病机制中起着重要作用。一方面,APOE的等位基因,尤其是APOEɛ4,可促进胶质细胞的炎症反应和促炎因子的分泌,引起神经炎症;另一方面,APOE可直接与Aβ相互作用,促进斑块沉积,oAβ的形成,同时降低Aβ的清除率。此外,APOE4在受损神经元中过度表达,可促进tau磷酸化以及tau介导的神经元死亡。APOE还可以通过诱导补体过度激活导致突触丢失,引起线粒体功能障碍,血脑屏障通透性增加等方式调控AD病理。近期的研究发现,微生物菌群对神经炎症的调节也通过APOE依赖的方式进行。除此之外,受到APOE的一些罕见的保护性突变的的启发,研究人员发现靶向APOE聚集的化合物可能会复制APOE3-Jacksonville变体的保护作用[21]。有关研究人员也对APOE3-Christchurch(R136S)做了深入的研究,利用tau病理小鼠模型和携带人APOE4的人iPSC诱导分化的神经元模型,含有纯合或杂合R136S突变。其中纯合的R136S突变挽救了APOE4驱动的tau病理、神经变性病变和神经炎症。杂合R136S突变能在一定程度上保护免受APOE4驱动的神经变性病变和神经炎症,但不能改善tau病理。同时单核RNA测序显示APOE4-R136S突变以基因剂量依赖的方式增加了疾病的保护性并减少了疾病相关细胞群[90]。
尽管淀粉样蛋白斑块和神经原纤维缠结被认为是AD 的主要病理特征,炎症和神经变性疾病之间也存在着恶性循环,但是用于研究AD 的小鼠模型并不能再现完整的神经病理学特征,导致AD 的治疗研究进展缓慢。AD 的诊治目前仍然存在未能有效满足临床需要。不过近年来国际上两种Aβ单抗治疗药物 - Aducanumab和Lecanemab的获批,使AD治疗领域的研究推进一大步。尤其是新进获批上市的靶向可溶性原纤维状Aβ的单抗Lecanemab(商品名,LeqembiTM),其三期临床试验结果显示,在为期18个月的双盲、安慰剂对照试验中,每两周对脑Aβ升高、轻度认知障碍或轻度痴呆的患者静脉注射一次,Lecanemab可将认知障碍和认知功能的恶化延迟约5个月[91-92]。除此之外,近期公布了三期临床试验结果的另一种靶向N3pG 修饰化的Aβ斑块单抗Donanemab,显示了其更加优良的治疗作用。与给与安慰剂的对照组相比,Donanamab单抗治疗将使得病人认知功能下降速度减缓了35%。在为期一年的三期临床试验中,近一半的实验组病人没有临床进展[93]。APOE调控AD的病理,在AD的发生发展中有着不可或缺的作用,并且在疾病的治疗过程中,APOE亚型也是一个必须要考虑的因素。在轻度认知障碍患者接受靶向Aβ的抗体治疗过程中,携带APOE4患者的治疗效果相对较差,并且更易罹患脑水肿[94]。因此,在使用靶向Aβ或者tau的药物的同时,联合使用针对APOE的治疗手段,也许能达到最佳的防治效果。
由于APOEɛ2在AD具有保护性作用,因此有观点认为如果能够在APOEɛ4携带者大脑中过表达APOE2,也许能对AD的发生和发展产生抑制作用。已有研究表明,在Aβ小鼠模型中过表达APOE2,可以有效降低Aβ斑块病理症状[95]。基于此发现,一项旨在APOEɛ4携带者大脑中,通过腺相关病毒载体(Adeno-associated virus,AAV)过表达APOE2的临床试验已经展开(NCT03634007)。然而,虽然过表达APOE2可以有效降低Aβ斑块的形成,但是在tau蛋白病理进程中,APOE2的过表达可能会加剧tau蛋白的磷酸化水平[39,96]。因此,过表达APOE2这项措施可能需要在AD疾病进程之初,即tau蛋白的病理还未起主导作用之前就开始实施,这对AD疾病的早期诊断提出了巨大的考验。此外,AAV病毒使用的剂量也是需要谨慎研究的内容。AAV的实际使用剂量需要使得病毒尽可能多的抵达AD影响的各个区域,同时,APOE2过表达的水平不能够破坏大脑中脂质代谢的平衡。
此外,与靶向Aβ的免疫抗体治疗类似,一些旨在降低大脑中APOE水平的抗体已经被开发[97]。然而,单纯降低APOE的水平也许很难达到有效治疗AD的目的。一方面,为了有效抑制Aβ斑块的形成,APOE水平的降低需要在Aβ病理症状形成之前就已经开始;另一方面,作为大脑中最主要的载脂蛋白,APOE水平的降低有可能会导致中枢神经系统脂质代谢的紊乱。基于上述的担忧,一种新型的靶向非脂质化APOE的抗体已经在多种AD小鼠模型中进行了测试[98-99]。在大脑中,非脂质化的APOE仅存在于Aβ斑块和血管沉积的致密核心处,因此,该抗体的使用在有效降低Aβ斑块和血管沉积数量的同时,并不会改变APOE总的含量。
目前,以APOE为靶点的治疗措施的开发尚处于早期阶段,针对APOE自身、APOE的细胞表面受体以及其保护性突变等为靶点的治疗方法的探索,可能为AD的治疗提供崭新的思路,最终使更多的AD患者受益。
| [1] | Yamazaki Y, Zhao N, Caulfield TR, et al. Apolipoprotein E and Alzheimer disease: pathobiology and targeting strategies[J]. Nat rev Neurol, 2019, 15(9):501-518. 本文引用 [1] 摘要 |
| [2] | Guo T, Zhang D, Zeng Y, et al. Molecular and cellular mechanisms underlying the pathogenesis of Alzheimer's disease[J]. Molecul neurodegener, 2020, 15(1):40. |
| [3] | Jansen IE, Savage JE, Watanabe K, et al. Genome-wide meta-analysis identifies new loci and functional pathways influencing Alzheimer's disease risk[J]. Nat Genet, 2019, 51(3):404-413. 本文引用 [1] 摘要 |
| [4] | Kunkle BW, Grenier-Boley B, Sims R, et al. Genetic meta-analysis of diagnosed Alzheimer's disease identifies new risk loci and implicates Aβ, tau, immunity and lipid processing[J]. Nat Genet, 2019, 51(3):414-430. 本文引用 [2] 摘要 |
| [5] | Witoelar A, Rongve A, Almdahl IS, et al. Meta-analysis of Alzheimer's disease on 9,751 samples from Norway and IGAP study identifies four risk loci[J]. Sci Rep, 2018, 8(1):18088. 本文引用 [1] 摘要 |
| [6] | Husain MA, Laurent B, Plourde M, et al. APOE and Alzheimer's Disease: From Lipid Transport to Physiopathology and Therapeutics[J]. Front Neurosci, 2021, 15:630502. 本文引用 [1] 摘要 |
| [7] | McIntosh AM, Bennett C, Dickson D, et al. The apolipoprotein E (APOE) gene appears functionally monomorphic in chimpanzees (Pan troglodytes)[J]. Plos One, 2012, 7(10):e47760. |
| [8] | Li Z, Shue F, Zhao N, et al. APOE2: protective mechanism and therapeutic implications for Alzheimer's disease[J]. Molecul Neurodegener, 2020, 15(1):63. |
| [9] | Hauser PS, Ryan RO. Impact of apolipoprotein E on Alzheimer's disease[J]. Current Alzheimer Res, 2013, 10(8):809-817. |
| [10] | Egert S, Rimbach G, Huebbe P, et al. ApoE genotype: from geographic distribution to function and responsiveness to dietary factors[J]. Proc Nutr Soc, 2012, 71(3):410-424. 本文引用 [1] 摘要 |
| [11] | Bras J, Guerreiro R, Darwent L, et al. Genetic analysis implicates APOE, SNCA and suggests lysosomal dysfunction in the etiology of dementia with Lewy bodies[J]. Hum Molecul Genet, 2014, 23(23):6139-6146. |
| [12] | Dhana K, Aggarwal NT, Rajan KB, et al. Impact of the Apolipoprotein E ε4 Allele on the Relationship Between Healthy Lifestyle and Cognitive Decline: A Population-Based Study[J]. Am J Epidemiol, 2021, 190(7):1225-1233. 本文引用 [1] 摘要 |
| [13] | Guerreiro R, Ross OA, Kun-Rodrigues C, et al. Investigating the genetic architecture of dementia with Lewy bodies: a two-stage genome-wide association study[J]. Lanc Neurol, 2018, 17(1):64-74. |
| [14] | Wennberg AM, Tosakulwong N, Lesnick TG, et al. Association of Apolipoprotein E ε4 With Transactive Response DNA-Binding Protein 43[J]. Jama Neurol, 2018, 75(11):1347-1354. 本文引用 [1] 摘要 |
| [15] | Yang HS, Yu L, White CC, et al. Evaluation of TDP-43 proteinopathy and hippocampal sclerosis in relation to APOE ε4 haplotype status: a community-based cohort study[J]. Lanc Neurol, 2018, 17(9):773-781. |
| [16] | Holtzman DM, Herz J, Bu G, et al. Apolipoprotein E and apolipoprotein E receptors: normal biology and roles in Alzheimer disease[J]. Cold Spring Harb Perspect Med, 2012, 2(3):a006312. |
| [17] | Shinohara M, Tachibana M, Kanekiyo T, et al. Role of LRP1 in the pathogenesis of Alzheimer's disease: evidence from clinical and preclinical studies[J]. J Lipid Res, 2017, 58(7):1267-1281. 本文引用 [1] 摘要 |
| [18] | Hu ML, Quinn J, Xue K, et al. Interactions between Apolipoprotein E Metabolism and Retinal Inflammation in Age-Related Macular Degeneration[J]. Life (Basel), 2021, 11(7). |
| [19] | Hatters DM, Peters-Libeu CA, Weisgraber KH, et al. Apolipoprotein E structure: insights into function[J]. Trends Biochem Sci, 2006, 31(8):445-454. 本文引用 [1] 摘要 |
| [20] | Le Guen Y, Raulin AC, Logue MW, et al. Association of African Ancestry-Specific APOE Missense Variant R145C With Risk of Alzheimer Disease[J]. Jama, 2023, 329(7):551-560. 本文引用 [1] 摘要 |
| [21] | Liu CC, Murray ME, Li X, et al. APOE3-Jacksonville (V236E) variant reduces self-aggregation and risk of dementia[J]. Sci Translat Med, 2021, 13(613):eabc9375. 本文引用 [2] 摘要 |
| [22] | Arboleda-Velasquez JF, Lopera F, O'Hare M, et al. Resistance to autosomal dominant Alzheimer's disease in an APOE3 Christchurch homozygote: a case report[J]. Nat Med, 2019, 25(11):1680-1683. 本文引用 [1] 摘要 |
| [23] | Ghiso J, Matsubara E, Koudinov A, et al. The cerebrospinal-fluid soluble form of Alzheimer's amyloid beta is complexed to SP-40,40 (apolipoprotein J), an inhibitor of the complement membrane-attack complex[J]. Biochem J, 1993, 293 (1):27-30. 本文引用 [1] 摘要 |
| [24] | Koldamova RP, Lefterov IM, Lefterova MI, et al. Apolipoprotein A-I directly interacts with amyloid precursor protein and inhibits A beta aggregation and toxicity[J]. Biochem, 2001, 40(12):3553-3560. |
| [25] | LaDu MJ, Falduto MT, Manelli AM, et al. Isoform-specific binding of apolipoprotein E to beta-amyloid[J]. J Biolog Chem, 1994, 269(38):23403-23406. |
| [26] | Manelli AM, Stine WB, Van Eldik LJ, et al. ApoE and Abeta1-42 interactions: effects of isoform and conformation on structure and function[J]. J Mol Neurosci, 2004, 23(3):235-246. 本文引用 [1] 摘要 |
| [27] | Wisniewski T, Golabek A, Matsubara E, et al. Apolipoprotein E: binding to soluble Alzheimer's beta-amyloid[J]. Biochem Biophys Res Commun, 1993, 192(2):359-365. |
| [28] | Tachibana M, Holm ML, Liu CC, et al. APOE4-mediated amyloid-β pathology depends on its neuronal receptor LRP1[J]. J Clin Invest, 2019, 129(3):1272-1277. 本文引用 [1] 摘要 |
| [29] | Verghese PB, Castellano JM, Garai K, et al. ApoE influences amyloid-β (Aβ) clearance despite minimal apoE/Aβ association in physiological conditions[J]. Proc National Acad Sci U S A, 2013, 110(19):E1807-E1816. |
| [30] | van Harten AC, Jongbloed W, Teunissen CE, et al. CSF ApoE predicts clinical progression in nondemented APOEε4 carriers[J]. Neurobiol Aging, 2017, 57:186-194. 本文引用 [1] 摘要 |
| [31] | Kloske CM, Wilcock DM. The Important Interface Between Apolipoprotein E and Neuroinflammation in Alzheimer's Disease[J]. Front Immunol, 2020, 11:754. 本文引用 [2] 摘要 |
| [32] | Mahley RW, Huang Y. Apolipoprotein e sets the stage: response to injury triggers neuropathology[J]. Neuron, 2012, 76(5):871-885. 本文引用 [1] 摘要 |
| [33] | Chen HK, Ji ZS, Dodson SE, et al. Apolipoprotein E4 domain interaction mediates detrimental effects on mitochondria and is a potential therapeutic target for Alzheimer disease[J]. J Biol Chem, 2011, 286(7):5215-5221. |
| [34] | Chernick D, Ortiz-Valle S, Jeong A, et al. Peripheral versus central nervous system APOE in Alzheimer's disease: Interplay across the blood-brain barrier[J]. Neurosci Lett, 2019, 708:134306. |
| [35] | Caberlotto L, Marchetti L, Lauria M, et al. Integration of transcriptomic and genomic data suggests candidate mechanisms for APOE4-mediated pathogenic action in Alzheimer's disease[J]. Sci Rep, 2016, 6:32583. 本文引用 [1] 摘要 |
| [36] | Thangavel R, Bhagavan SM, Ramaswamy SB, et al. Co-Expression of Glia Maturation Factor and Apolipoprotein E4 in Alzheimer's Disease Brain[J]. J Alzheimers Diseas, 2018, 61(2):553-560. |
| [37] | Tulloch J, Leong L, Thomson Z, et al. Glia-specific APOE epigenetic changes in the Alzheimer's disease brain[J]. Brain Res, 2018, 1698:179-186. 本文引用 [1] 摘要 |
| [38] | Rodriguez GA, Tai LM, LaDu MJ, et al. Human APOE4 increases microglia reactivity at Aβ plaques in a mouse model of Aβ deposition[J]. J Neuroinflammation, 2014, 11:111. |
| [39] | Shi Y, Yamada K, Liddelow SA, et al. ApoE4 markedly exacerbates tau-mediated neurodegeneration in a mouse model of tauopathy[J]. Nat, 2017, 549(7673):523-527. |
| [40] | Maezawa I, Nivison M, Montine KS, et al. Neurotoxicity from innate immune response is greatest with targeted replacement of E4 allele of apolipoprotein E gene and is mediated by microglial p38MAPK[J]. FASEB J, 2006, 20(6):797-799. 本文引用 [1] 摘要 |
| [41] | Ulrich JD, Ulland TK, Mahan TE, et al. ApoE facilitates the microglial response to amyloid plaque pathology[J]. J Experiment Med, 2018, 215(4):1047-1058. 本文引用 [2] 摘要 |
| [42] | Hickman SE, Allison EK, El Khoury J, et al. Microglial dysfunction and defective beta-amyloid clearance pathways in aging Alzheimer's disease mice[J]. J Neurosci, 2008, 28(33):8354-8360. 本文引用 [1] 摘要 |
| [43] | Anderson MA, Burda JE, Ren Y, et al. Astrocyte scar formation aids central nervous system axon regeneration[J]. Nat, 2016, 532(7598):195-200. |
| [44] | Lin YT, Seo J, Gao F, et al. APOE4 Causes Widespread Molecular and Cellular Alterations Associated with Alzheimer's Disease Phenotypes in Human iPSC-Derived Brain Cell Types[J]. Neuron, 2018, 98(6):1294. |
| [45] | Zhao J, Davis MD, Martens YA, et al. APOE ε4/ε4 diminishes neurotrophic function of human iPSC-derived astrocytes[J]. Hum Mol Genet, 2017, 26(14):2690-2700. 本文引用 [1] 摘要 |
| [46] | Chung WS, Verghese PB, Chakraborty C, et al. Novel allele-dependent role for APOE in controlling the rate of synapse pruning by astrocytes[J]. Proc National Acad Sci U S A, 2016, 113(36):10186-10191. 本文引用 [1] 摘要 |
| [47] | Larramona-Arcas R, González-Arias C, Perea G, et al. Sex-dependent calcium hyperactivity due to lysosomal-related dysfunction in astrocytes from APOE4 versus APOE3 gene targeted replacement mice[J]. Molecul Neurodegener, 2020, 15(1):35. |
| [48] | Kang SS, Ebbert MTW, Baker KE, et al. Microglial translational profiling reveals a convergent APOE pathway from aging, amyloid, and tau[J]. J Exp Med, 2018, 215(9):2235-2245. 本文引用 [1] 摘要 |
| [49] | Rebeck GW, Reiter JS, Strickland DK, et al. Apolipoprotein E in sporadic Alzheimer's disease: allelic variation and receptor interactions[J]. Neuron, 1993, 11(4):575-580. 本文引用 [1] 摘要 |
| [50] | Youmans KL, Tai LM, Nwabuisi-Heath E, et al. APOE4-specific changes in Aβ accumulation in a new transgenic mouse model of Alzheimer disease[J]. J Biol Chem, 2012, 287(50):41774-41786. 本文引用 [1] 摘要 |
| [51] | Gratuze M, Jiang H, Wang C, et al. APOE Antibody Inhibits Aβ-Associated Tau Seeding and Spreading in a Mouse Model[J]. Ann Neurol, 2022, 91(6):847-852. 本文引用 [1] 摘要 |
| [52] | Mahan TE, Wang C, Bao X, et al. Selective reduction of astrocyte apoE3 and apoE4 strongly reduces Aβ accumulation and plaque-related pathology in a mouse model of amyloidosis[J]. Mol Neurodegener, 2022, 17(1):13. 本文引用 [1] 摘要 |
| [53] | Koffie RM, Hashimoto T, Tai HC, et al. Apolipoprotein E4 effects in Alzheimer's disease are mediated by synaptotoxic oligomeric amyloid-β[J]. Brain, 2012, 135(7):2155-2168. |
| [54] | Liu CC, Hu J, Zhao N, et al. Astrocytic LRP1 Mediates Brain Aβ Clearance and Impacts Amyloid Deposition[J]. J Neurosci, 2017, 37(15):4023-4031. 本文引用 [1] 摘要 |
| [55] | Ma Q, Zhao Z, Sagare AP, et al. Blood-brain barrier-associated pericytes internalize and clear aggregated amyloid-β42 by LRP1-dependent apolipoprotein E isoform-specific mechanism[J]. Mol Neurodegener, 2018, 13(1):57. 本文引用 [2] 摘要 |
| [56] | Hori Y, Hashimoto T, Nomoto H, et al. Role of Apolipoprotein E in β-Amyloidogenesis: Isoform-specific effects on protofibril to fibril conversion of Aβ in vitro and brain Aβ deposition in vivo[J]. J Biol Chem, 2015, 290(24):15163-15174. 本文引用 [1] 摘要 |
| [57] | Liu CC, Zhao N, Fu Y, et al. ApoE4 Accelerates Early Seeding of Amyloid Pathology[J]. Neuron, 2017, 96(5):1024-1032.e3. 本文引用 [1] 摘要 |
| [58] | Jiang Q, Lee CY, Mandrekar S, et al. ApoE promotes the proteolytic degradation of Abeta[J]. Neuron, 2008, 58(5):681-693. 本文引用 [1] 摘要 |
| [59] | Muth C, Hartmann A, Sepulveda-Falla D, et al. Phagocytosis of Apoptotic Cells Is Specifically Upregulated in ApoE4 Expressing Microglia in vitro[J]. Front Cell Neurosci, 2019, 13:181. 本文引用 [1] 摘要 |
| [60] | Hou X, Zhang X, Zou H, et al. Differential and substrate-specific inhibition of γ-secretase by the C-terminal region of ApoE2, ApoE3, and ApoE4[J]. Neuron, 2023, 111(12):1989-1913.e5. |
| [61] | Wang C, Xiong M, Gratuze M, et al. Selective removal of astrocytic APOE4 strongly protects against tau-mediated neurodegeneration and decreases synaptic phagocytosis by microglia[J]. Neuron, 2021, 109(10):1657-1674.e7. 本文引用 [1] 摘要 |
| [62] | Brecht WJ, Harris FM, Chang S, et al. Neuron-specific apolipoprotein e4 proteolysis is associated with increased tau phosphorylation in brains of transgenic mice[J]. J Neurosci, 2004, 24(10):2527-2534. 本文引用 [1] 摘要 |
| [63] | Koutsodendris N, Blumenfeld J, Agrawal A, et al. Neuronal APOE4 removal protects against tau-mediated gliosis, neurodegeneration and myelin deficits[J]. Nat Aging, 2023, 3(3):275-296. 本文引用 [2] 摘要 |
| [64] | Andrews-Zwilling Y, Bien-Ly N, Xu Q, et al. Apolipoprotein E4 causes age- and Tau-dependent impairment of GABAergic interneurons, leading to learning and memory deficits in mice[J]. J Neurosci, 2010, 30(41):13707-13717. 本文引用 [1] 摘要 |
| [65] | Najm R, Jones EA, Huang Y, et al. Apolipoprotein E4, inhibitory network dysfunction, and Alzheimer's disease[J]. Mol Neurodegener, 2019, 14(1):24. 本文引用 [1] 摘要 |
| [66] | Gomez-Arboledas A, Davila JC, Sanchez-Mejias E, et al. Phagocytic clearance of presynaptic dystrophies by reactive astrocytes in Alzheimer's disease[J]. Glia, 2018, 66(3):637-653. 本文引用 [1] 摘要 |
| [67] | Hong S, Beja-Glasser VF, Nfonoyim BM, et al. Complement and microglia mediate early synapse loss in Alzheimer mouse models[J]. Sci, 2016, 352(6286):712-716. |
| [68] | McGeer PL, Walker DG, Pitas RE, et al. Apolipoprotein E4 (ApoE4) but not ApoE3 or ApoE2 potentiates beta-amyloid protein activation of complement in vitro[J]. Brain Res, 1997, 749(1):135-138. 本文引用 [1] 摘要 |
| [69] | Wang W, Zhao F, Ma X, et al. Mitochondria dysfunction in the pathogenesis of Alzheimer's disease: recent advances[J]. Mol Neurodegene, 2020, 15(1):30. 本文引用 [1] 摘要 |
| [70] | Mahley RW, Weisgraber KH, Huang Y, et al. Apolipoprotein E4: a causative factor and therapeutic target in neuropathology, including Alzheimer's disease[J]. Proc Nat Acad Sci U S A, 2006, 103(15):5644-5651. 本文引用 [1] 摘要 |
| [71] | Valla J, Yaari R, Wolf AB, et al. Reduced posterior cingulate mitochondrial activity in expired young adult carriers of the APOE ε4 allele, the major late-onset alzheimer's susceptibility gene[J]. J Alzheimers Disease, 2010, 22(1):307-313. |
| [72] | Ihara Y, Hayabara T, Sasaki K, et al. Relationship between oxidative stress and apoE phenotype in Alzheimer's disease[J]. Acta neurol Scand, 2000, 102(6):346-349. 本文引用 [1] 摘要 |
| [73] | Robb WH, Khan OA, Ahmed HA, et al. Lower cerebral oxygen utilization is associated with Alzheimer's disease-related neurodegeneration and poorer cognitive performance among apolipoprotein E ε4 carriers[J]. J Cereb Blood Flow Metab, 2022, 42(4):642-655. 本文引用 [1] 摘要 |
| [74] | Sun YY, Wang Z, Huang HC, et al. Roles of ApoE4 on the Pathogenesis in Alzheimer's Disease and the Potential Therapeutic Approaches[J]. Cell Molr Neurobiol, 2023,1-22. |
| [75] | Sperling R, Salloway S, Brooks DJ, et al. Amyloid-related imaging abnormalities in patients with Alzheimer's disease treated with bapineuzumab: a retrospective analysis[J]. Lanc Neurol, 2012, 11(3):241-249. |
| [76] | Yamazaki Y, Shinohara M, Yamazaki A, et al. ApoE (Apolipoprotein E) in Brain Pericytes Regulates Endothelial Function in an Isoform-Dependent Manner by Modulating Basement Membrane Components[J]. Arterioscler Thromb Vasc Biol, 2020, 40(1):128-144. 本文引用 [1] 摘要 |
| [77] | Sakai K, Boche D, Carare R, et al. Aβ immunotherapy for Alzheimer's disease: effects on apoE and cerebral vasculopathy[J]. Acta neuropathologica, 2014, 128(6):777-789. 本文引用 [1] 摘要 |
| [78] | Sagare AP, Bell RD, Zhao Z, et al. Pericyte loss influences Alzheimer-like neurodegeneration in mice[J]. Nature communicat, 2013, 4:2932. |
| [79] | Blanchard JW, Bula M, Davila-Velderrain J, et al. Reconstruction of the human blood-brain barrier in vitro reveals a pathogenic mechanism of APOE4 in pericytes[J]. Nat Med, 2020, 26(6):952-963. 本文引用 [1] 摘要 |
| [80] | Atagi Y, Liu CC, Painter MM, et al. Apolipoprotein E Is a Ligand for Triggering Receptor Expressed on Myeloid Cells 2 (TREM2)[J]. J Biol Chem, 2015, 290(43):26043-26050. 本文引用 [1] 摘要 |
| [81] | Hickman SE, Kingery ND, Ohsumi TK, et al. The microglial sensome revealed by direct RNA sequencing[J]. Nat Neurosci, 2013, 16(12):1896-1905. 本文引用 [1] 摘要 |
| [82] | Kleinberger G, Yamanishi Y, Suárez-Calvet M, et al. TREM2 mutations implicated in neurodegeneration impair cell surface transport and phagocytosis[J]. Sci Transl Med, 2014, 6(243):243ra86. |
| [83] | Colonna M, Wang Y. TREM 2 variants: new keys to decipher Alzheimer disease pathogenesis[J]. Nat Rev Neurosci, 2016, 17(4):201-207. 本文引用 [1] 摘要 |
| [84] | Ma LZ, Tan L, Bi YL, et al. Dynamic changes of CSF sTREM2 in preclinical Alzheimer's disease: the CABLE study[J]. Mol Neurodegener, 2020, 15(1):25. 本文引用 [1] 摘要 |
| [85] | Suárez-Calvet M, Morenas-Rodríguez E, Kleinberger G, et al. Early increase of CSF sTREM2 in Alzheimer's disease is associated with tau related-neurodegeneration but not with amyloid-β pathology[J]. Mol Neurodegener, 2019, 14(1):1. 本文引用 [1] 摘要 |
| [86] | Tzioras M, Davies C, Newman A, et al. Invited Review: APOE at the interface of inflammation, neurodegeneration and pathological protein spread in Alzheimer's disease[J]. Neuropathol Appl Neurobiol, 2019, 45(4):327-346. |
| [87] | Seo DO, O'Donnell D, Jain N, et al. ApoE isoform- and microbiota-dependent progression of neurodegeneration in a mouse model of tauopathy[J]. Sci(NY), 2023, 379(6628):eadd1236. |
| [88] | Sun Y, Baptista LC, Roberts LM, et al. The Gut Microbiome as a Therapeutic Target for Cognitive Impairment[J]. J Gerontol A Biol Sci Med Sci, 2020, 75(7):1242-1250. 摘要 |
| [89] | Seo DO, Boros BD, Holtzman DM, et al. The microbiome: A target for Alzheimer disease[J]. Cell Res, 2019, 29(10):779-780. |
| [90] | Nelson M. The APOE4-R136S mutation protects against APOE4-driven Tau pathology[J]. Neurodegener Neuroinflammat, 2023. |
| [91] | Söderberg L, Johannesson M, Nygren P, et al. Lecanemab, Aducanumab, and Gantenerumab - Binding Profiles to Different Forms of Amyloid-Beta Might Explain Efficacy and Side Effects in Clinical Trials for Alzheimer's Disease[J]. Neurotherapeutics, 2023, 20(1):195-206. 本文引用 [1] 摘要 |
| [92] | Knopman DS, Hershey L. Implications of the Approval of Lecanemab for Alzheimer Disease Patient Care: Incremental Step or Paradigm Shift[J]. Neurol, 2023, 10.1212/WNL.000000000020 7438. |
| [93] | Bateman RJ, Cummings J, Schobel S, et al. Lilly's Donanemab Significantly Slowed Cognitive and Functional Decline in Phase 3 Study of Early Alzheimer's Disease[J]. Alzheimers Res Ther, 2022, 14(1):178. 本文引用 [1] 摘要 |
| [94] | Kennedy RE, Cutter GR, Schneider LS, et al. Effect of APOE genotype status on targeted clinical trials outcomes and efficiency in dementia and mild cognitive impairment resulting from Alzheimer's disease[J]. Alzheimers Dement, 2014, 10(3):349-359. 本文引用 [1] 摘要 |
| [95] | Zhao L, Gottesdiener AJ, Parmar M, et al. Intracerebral adeno-associated virus gene delivery of apolipoprotein E2 markedly reduces brain amyloid pathology in Alzheimer's disease mouse models[J]. Neurobiol Aging, 2016, 44:159-172. 本文引用 [1] 摘要 |
| [96] | Zhao N, Liu CC, Van Ingelgom AJ, et al. APOE ε2 is associated with increased tau pathology in primary tauopathy[J]. Nat Commun, 2018, 9(1):4388. 本文引用 [1] 摘要 |
| [97] | Kim J, Eltorai AE, Jiang H, et al. Anti-apoE immunotherapy inhibits amyloid accumulation in a transgenic mouse model of Aβ amyloidosis[J]. J Experiment Med, 2012, 209(12):2149-2156. 本文引用 [1] 摘要 |
| [98] | Liao F, Li A, Xiong M, et al. Targeting of nonlipidated, aggregated apoE with antibodies inhibits amyloid accumulation[J]. J Clin Invest, 2018, 128(5):2144-2155. 本文引用 [1] 摘要 |
| [99] | Xiong M, Jiang H, Serrano JR, et al. APOE immunotherapy reduces cerebral amyloid angiopathy and amyloid plaques while improving cerebrovascular function[J]. Sci Translat Med, 2021, 13(581). |