缺血性脑卒中往往因为缺血缺氧,导致缺血半暗带神经元变性坏死
[1],“治疗性血管新生”成为国内外治疗缺血性脑卒中的研究热点
[2],通过促进治疗性血管新生,进而增加血供,达到挽救休眠神经元的目的
[3]。当血管内皮细胞处于重度缺氧的环境下,自噬会抑制血管新生的发生
[4]。研究发现PI3K/Akt/mTOR通路是挽救脑细胞,恢复神经功能的潜在靶点,是参与内皮细胞形成新生血管的关键性靶点,另据表明,该通路调控血管新生主要与抑制自噬和凋亡有关
[5]。黄芪为豆科植物属蒙古黄芪、膜荚黄芪的干燥根,是历史悠久、应用广泛的传统补益中药
[6]。黄芪的主要成分有黄芪多糖、黄芪皂苷、氨基酸等。黄芪多糖(Astragalus polysaccharide,APS)是黄芪的主要活性成分,具有广泛的药理活性和作用机制,可促进血管内皮细胞增殖,达到促血管新生的目的
[7⇓-9],可抑制急性脑缺血再灌注损伤诱导的细胞自噬
[10]。近年来有研究发现,黄芪多糖对脑缺血损伤有保护作用,但其改善脑缺血与促进脑血管新生的相关性及自噬作用机制鲜有报道。本研究采用血管段三维培养和缺氧条件下体外内皮细胞培养模型,研究黄芪多糖促进血管新生机制与自噬靶点的关系,为该药在今后的药品开发中提供实验依据。
1 材料与方法
1.1 实验材料
1.1.1 实验动物
2月龄雄性SD大鼠,体质量约(200±20)g,均购于中国医学科学院医学实验动物研究所,动物生产许可证号SCXK(京)20140004,本研究所设计的动物实验符合中国伦理委员会相关动物研究指导原则(编号TCMLAEC2012011)。
1.1.2 实验细胞
大鼠脑微血管内皮细胞(RBMEC,商城北纳创联生物科技有限公司,批次:BNCC337717)
1.1.3 仪器设备
DM3000B型倒置显微镜(德国Leica公司);ENSPINE型多功能读板机(美国 Perkin Elmer公司);Fusion FX5型化学发光凝胶成像系统(法国Vilber公司);min-subcellget型垂直电泳槽(美国 Bio-rad公司);Mini 型转膜仪(中国北京凯元仪器公司);CDI-165M 型三气培养箱(日本 Astec 公司);DL-CJ-2NDI 型超净台(中国北京东联哈尔仪器制造有限公司);ALLEGRA 61R型低温高速冷冻离心机(美国 Beckman 公司);Eppendorf Reference® 2微量移液器(德国 Eppendorf 公司)。
1.1.4 实验试剂
鼠尾胶(美国 Sigma 公司,货号:5056-A);MCDB131,DMEM 培养基(美国 Gibco 公司,批号:10372019,12430054);特级胎牛血清(美国 Gibco 公司,货号:10091148);D-hanks 缓冲液(美国Gibco公司,货号:14025092);BCA蛋白浓度测定试剂盒(北京索莱宝科技有限公司,货号:PC0020);trizol(北京索莱宝科技有限公司,货号:15596026);甘油醛3-磷酸脱氢酶(GAPDH,美国 Abcam 公司,批号:ab209344);PBS 磷酸盐缓冲液干粉(北京索莱宝科技有限公司,货号:P1010);胰蛋白酶-EDTA溶液(北京索莱宝科技有限公司,货号:T1300);CCK-8试剂盒(上海尚宝生物科技有限公司,货号:ST1006);LC3B、Becline-1、GAPDH抗体(美国CST公司,货号分别为;3868、3495、5174);P62抗体(美国CST公司,货号:5114S);SDS-PAGE 4x上样缓冲液(北京索莱宝科技有限公司,货号:P1015);20x TBS(北京索莱宝科技有限公司,货号:T1080);羊抗兔二抗(美国 Abcam 公司,货号:ab6795);高灵敏ECL发光试剂盒(北京索莱宝科技有限公司,货号:PE0010);黄芪多糖冻干粉(北京索莱宝科技有限公司,货号:SA9790,纯度 ≥ 98 %);CoCl2(北京索莱宝科技有限公司,货号:H9060);雷帕霉素(索莱宝科技有限公司,货号:R8140);MEM 培养基(美国 Gibco公司,货号:41090036)。巴弗洛霉素A1(美国Med Chem Express公司,货号:HY-100558)。
1.2 实验方法
1.2.1 三维培养血管新生模型
根据ZHU WH的研究构建三维培养血管新生模型
[11]。首先将大鼠颈椎脱臼处死,浸没于75%乙醇中消毒。在超净台上迅速打开大鼠胸腔,取出1 cm胸主动脉,置于含预冷D-hanks的培养皿中,清洗2次,切取1 mm动脉环。在96孔板内铺好pH 7.4鼠尾胶,37 ℃凝固30 min后将动脉环置入,然后再加入等量鼠尾胶,再次凝固30 min后于上层每孔加入100 mL含MCDB131混合培养液。将含有血管段的培养板放入37 ℃,5% CO
2条件培养,2~3天更换一次培养基,观察血管出芽情况,用于后续实验检测。
1.2.2 细胞培养与传代
将购买的RBMEC种于75 cm2培养瓶中,内放细胞培养基15 mL(DMEM+10%胎牛血清+1%双抗),于37 ℃,5% CO2培养,每2~3天换液培养,待传2~3代后,内皮细胞经胰酶消化,分别种于96孔板和6孔培养板,以备实验检测使用。
1.2.3 缺氧模型制备
根据张迪等使用氯化钴造缺氧模型
[12],将细胞种于96孔板后,置于37 ℃和5% CO
2混合空气的恒温孵箱中常规培养24 h后,在相同气体培养条件下,更换培养基为2% 胎牛血清+高糖DMEM均一化24 h,再行缺氧模型制备。
本实验以CoCl2分别干预RBMEC制备化学缺氧模型,设立5个不同浓度的CoCl2缺氧组,分别为100、200、300、400 μmol·L-1、以及空白对照组。通过CCK-8法对不同浓度的CoCl2致RBMEC缺氧24 h后活性进行检测。旨在探讨经CoCl2缺氧制备周细胞缺氧模型的最佳CoCl2浓度(即CoCl2缺氧浓度)。根据张迪等研究结论,400 μmol·L-1CoCl2浓度能造成细胞重度缺氧环境,符合本文研究条件,预实验结果与其结论相一致,故选用400 μmol·L-1CoCl2浓度作为RBMEC制备缺氧模型的最佳CoCl2浓度。
1.2.4 给药剂量确定
配制1.0 g·L-1的APS母液,经滤器过滤后,制备浓度为10、25、45 mg·L-1的APS培养基。以每孔200 μL的剂量,将不同浓度APS培养基分别接种于孔板底部已铺满细胞的96孔板内,同时设置空白对照组(加入原培养基200 μL)培养24 h后,通过CCK-8法对不同浓度的APS促RBMEC增殖活性进行检测,最终确定25 mg·L-1浓度为本实验的给药剂量。
1.2.5 实验分组:
(1)按照上述方法进行新生血管模型制备后,随机分为2组,并进行给药干预:
对照组(Control):常规培养基 MCDB131
黄芪多糖组(APS):常规培养基 MCDB131+25 mg·L-1APS
以上各组均每三天换液,连续培养14天。
(2)对RBMEC增殖、自噬蛋白表达指标检测。随机分为五组:
正常对照组(Control):常规培养基+空白培养基
模型组(Model):常规培养基+400 μmol·L-1CoCl2
黄芪多糖组(APS):常规培养基+400 μmol·L-1CoCl2+25 mg·L-1APS
雷帕霉素组(RAPA):常规培养基+400 μmol·L-1CoCl2+10 mg·L-1RAPA
巴弗洛霉素组(BafA1):常规培养基+400 μmol·L-1CoCl2+20 mg·L-1BafA1
黄芪多糖和雷帕霉素组(APS+RAPA):常规培养基+400 μmol·L-1CoCl2+25 mg·L-1APS +10 mg·L-1RAPA
黄芪多糖和巴弗洛霉素组(APS+BafA1):常规培养基+400 μmol ·L-1CoCl2+25 mg·L-1APS+20 mg·L-1BafA1
1.3 检测指标
1.3.1 新生血管形态学观察
药物干预结束后,利用倒置显微镜观察各组胸主动脉血管段上的血管新生形态及数量,调节微调在不同层面,随机选取5个视野(100×)计数成管的新生血管数,评价黄芪多糖促血管新生情况。
1.3.2 CCK-8检测RBMEC增殖情况
RBMEC接种密度为1x105 个/mL,96孔板每孔接种100 μL,每组6个复孔。除正常培养RBMEC细胞外,其余各组弃掉原培养液后加入400 μmol·L-1 CoCl2,24 h后按组分别给予相应浓度的黄芪多糖、雷帕霉素等药物。再行24 h培养后,每孔加入CCK-8溶液10 μL(避光),于37 ℃孵育0.5 h、1 h、1.5 h、2 h,450 nm波长下,在酶联免疫检测仪上测定各孔的光吸收值。
1.3.3 Western blot法检测各组细胞中LC3B、Beclin-1、p62的蛋白表达
各组弃细胞培养液后用预冷PBS洗细胞3次,裂解细胞,离心提取总蛋白,利用BCA试剂盒对蛋白进行定量,均一化。按每孔5 μg总蛋白加样,各组逐一上样,电泳后转膜。PVDF膜条置于TBST液中,于摇床室温封闭2 h。后弃液加入相应一抗LC3B (1:1500)、p62 (1:1000)、Beclin-1(1:1000)及GAPDH(1:2000),4 ℃摇床过夜。再经TBST充分洗涤PVDF膜三次后加二抗,ab6721(1:10000)(羊抗兔来源),室温摇床孵育2 h。显影后利用凝胶成像系统拍照,并进行灰度信号分析,以GAPDH为内参照,计算LC3B、Beclin-1、p62蛋白相对表达量,每组重复测量3次,取其平均值。
1.3.4 统计学方法
通过SPSS21.0统计学软件进行处理。计量资料用均数±标准差(�¯±�)表示,多组间比较采用方差分析。以P<0.05为差异有统计学意义。
2 结果与分析
2.1 APS具有促进血管新生的作用
血管段实验为三维立体培养,利用倒置显微镜观察同一焦平面下培养2 d、6 d、10 d、13 d后内皮细胞与血管新生情况。各组实验结果显示:培养2 d后,观察到血管段附近出现内皮细胞沿条索状排列形成管状,存在新生血管出芽现象,各组无明显差异。6 d后,观察到各组新生血管增多,APS组新生血管数量明显多于对照组(P<0.01)。10 d后,各组健存新生血管呈放射管网状向血管段周围爬行,立体感强,呈现相对稳定、成熟状态(见图1),计数新生血管成管数量显示,与同一时间的对照组比较,APS组给予黄芪多糖,稳定存在的新生血管数量增多(P<0.05,见表1)。随着培养时间延长,13 d后各组新生血管均有退化现象出现,内皮细胞皱缩,细胞与细胞分离。
图1 黄芪多糖对胸主动脉血管段新生血管数量的影响(�¯±�,n=6)Fig1 Effect of astragalus polysaccharide on neovascularization of aortic segment (×100) Not: The blue arrow represents endothelial cells; The red arrow indicates neovascularization. |
Full size|PPT slide
表1 黄芪多糖对胸主动脉血管段新生血管数量的影响 (�¯±�,n=6)Tab 1 Effect of astragalus polysaccharide on the number of new vessels in aortic segment(�¯±�,n=6) |
| 组别 | 质量浓度 /mmol·L-1 | 培养2d后新生血管数(根) | 培养6d后新生血管数(根) | 培养10 d后新生血管数(根) | 培养13 d后新生血管数(根) |
|---|
| Control | 0.01 | 4.67±1.15 | 8.8±4.15 | 8.00±3.56 | 5.00±3.74 |
| APS | 0.10 | 11.5±0.71 | 11.67±4.1△△ | 13.80±2.68△ | 9.25±4.03△ |
| Not: Compared with the control group at the same time, △P<0.05,△△P<0.05 in each APS group |
2.2 APS具有促进内皮细胞增殖的作用
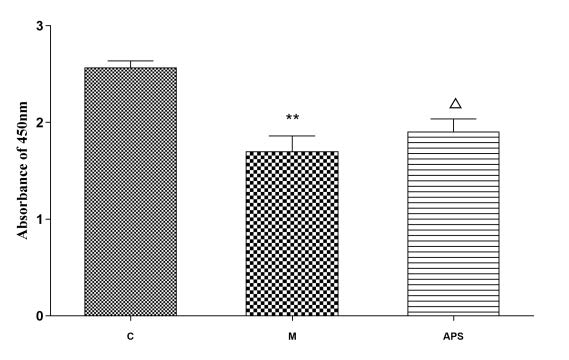
各组经不同条件干预后,实验结果显示:与对照组相比,在缺氧条件下,RBMEC增殖活力明显下降(P<0.01);与模型组相比,在缺氧条件下,APS可增强内皮细胞活性,促进内皮细胞增殖(P<0.05,见表2,图2)。
表2 缺氧条件下黄芪多糖对内皮细胞增殖的影响(�¯±�,n=6)Tab 2 Effects of astragalus polysaccharides on endothelial cell proliferation under hypoxia condition(�¯±�,n=6) |
| Group | n | 细胞活力(OD value) |
|---|
| Control | 6 | 2.57±0.0٦ |
| Model | 6 | 1.69±0.14** |
| APS | 6 | 1.91±0.12△ |
| Not: Compared with the Control group, **P<0.01; △P<0.05 compared to Model group |
图 2 缺氧条件下黄芪多糖对内皮细胞增殖的影响(�¯±�,n=6)Fig 2 Effects of astragalus polysaccharides on endothelial cell proliferation under hypoxia condition(�¯±�,n=6) |
Full size|PPT slide
2.3 APS具有下调自噬蛋白表达的作用
各组行不同条件干预后检测Becline-1、LC3B、p62蛋白表达情况,实验结果显示(见表3,图4):与对照组相比,缺氧条件下,Becline-1、LC3B蛋白表达明显上调(P<0.05),p62蛋白表达明显下调(P<0.01);经APS干预后Becline-1、LC3B蛋白表达量下调(P<0.01)。
表3 各组内皮细胞中Becline-1、LC3B、p62蛋白表达的检测结果(�¯±�,n=6)Tab 3 Expression of autophagy markers Becline-1、LC3B and p62 in each group(�¯±�,n=6) |
| Group | Becline-1 | LC3B | p62 |
|---|
| Control | 1.28±0.06 | 0.99±0.07 | 1.48±0.07 |
| Model | 1.90±0.09** | 1.39±0.10** | 1.26±0.03* |
| APS | 0.84±0.04△△ | 0.97±0.07△△ | 1.17±0.04 |
| Not: Compared with the Control group, * P<0.05, **P<0.01; Compared with the model group, △△ P<0.01. |
2.4 在自噬激动剂和抑制剂作用下,明确APS调控自噬靶点的作用
2.4.1 APS在自噬激动剂和抑制剂作用下对细胞的增殖作用
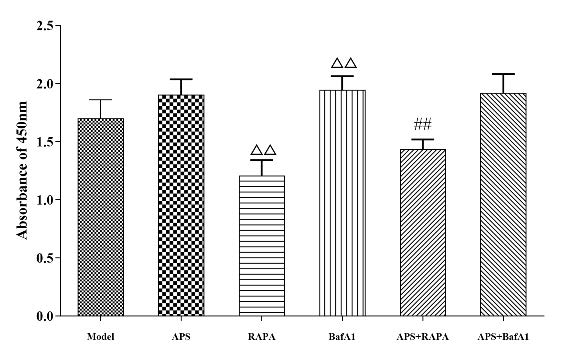
为了更好地明确APS调控自噬蛋白的靶点,行自噬激动剂和抑制剂干预后,细胞增殖实验结果显示:与Model组相比,经RAPA干预后细胞增殖活力明显减弱(P<0.01)。与RAPA组相比,经过APS干预后,细胞增殖活力明显增加(P<0.01),黄芪多糖与雷帕霉素形成拮抗关系。与Model组相比,BafA1细胞增殖活力明显增加(P<0.01),趋势与APS组相似。与BafA1组相比,经过APS干预后,细胞增殖活力未见明显差异。(表4,图3)。
表4 缺氧条件下黄芪多糖对内皮细胞增殖的影响(�¯±�,n=6)Tab 4 Effects of astragalus polysaccharides on endothelial cell proliferation under hypoxia condition(�¯±�,n=6) |
| Group | n | 细胞活力(OD value) |
|---|
| Model | 6 | 1.69±0.14 |
| APS | 6 | 1.91±0.12 |
| RAPA | 6 | 1.21±0.12△△ |
| BafA1 | 6 | 1.95±0.11△△ |
| APS+RAPA | 6 | 1.44±0.08## |
| APS+BafA1 | 6 | 1.92±0.15 |
| Not: Compared with Medol group, △△P<0.01; Compared with RAPA, ##P<0.01. |
图3 缺氧条件下黄芪多糖对内皮细胞增殖的影响(�¯±�,n=6)Fig 3 Effects of astragalus polysaccharides on endothelial cell proliferation under hypoxia condition(�¯±�,n=6) |
Full size|PPT slide
2.4.2 APS在自噬激动剂和抑制剂作用下,明确调控自噬蛋白靶点的作用
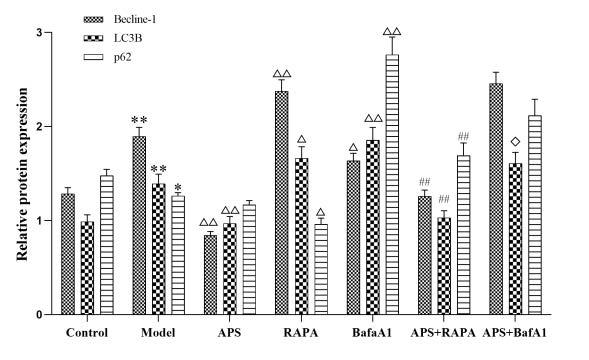
为了更好地明确APS调控自噬蛋白的靶点,行自噬激动剂和抑制剂干预后,蛋白印迹实验结果显示(表5,图4-5):
表 5 各组内皮细胞中Becline-1、LC3B、p62蛋白表达的检测结果(�¯±�,n=6))Tab 5 Expression of autophagy markers Becline-1、LC3B and p62 in each group(�¯±�,n=6) |
| Group | Becline-1 | LC3B | p62 |
|---|
| Control | 1.28±0.06 | 0.99±0.07 | 1.48±0.07 |
| Model | 1.90±0.09** | 1.39±0.10** | 1.26±0.03* |
| APS | 0.84±0.04△△ | 0.97±0.07△△ | 1.17±0.04 |
| RAPA | 2.38±0.12△△ | 1.66±0.12△ | 0.96±0.06△ |
| BafA1 | 1.63±0.08△ | 1.85±0.13△△ | 2.76±0.19△△ |
| APS+RAPA | 1.26±0.06## | 1.03±0.08## | 1.69±0.13## |
| APS+BafA1 | 2.46±0.11 | 1.61±0.12◇ | 2.11±0.17 |
| Not: Compared with the Control group, *P<0.05, **P<0.01; Compared with the model group, △P<0.05, △△P<0.01; Compared with RAPA group, ##P<0.05; Compared to BafA1, ◇P < 0.05. |
图4 各组内皮细胞中Becline-1、LC3B、p62蛋白表达的检测结果(�¯±�,n=3)Fig 4. Expression of autophagy markers Becline-1、LC3B and p62 in each group(�¯±�,n=3) |
Full size|PPT slide
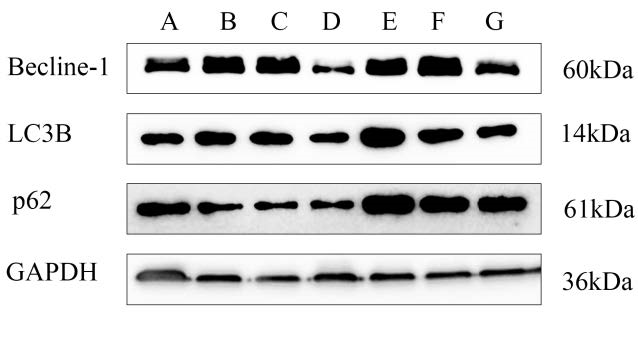
图 5 各组内皮细胞中Becline-1、LC3B、p62蛋白表达的检测结果(�¯±�,n=3)Fig 5. Expression of autophagy markers Becline-1、LC3B and p62 in each group(�¯±�,n=3) Not:A: Control Group; B:Model Group; C: RAPA Group; D:APS Group; E:BafA1 Group ; F: APS+ BafA1 Group; G: APS+RAPA Group |
Full size|PPT slide
1.经RAPA干预后,Becline-1、LC3B蛋白表达量明显增加(P<0.05,P<0.01),p62蛋白表达量明显减少(P<0.05),符合RAPA自噬激动剂蛋白表达变化规律。与RAPA组相比,经APS干预后Becline-1、LC3B蛋白表达量下调(P<0.01),p62蛋白表达明显上调(P<0.01)。
2.与M组相比,经BafA1干预后,Becline-1蛋白表达降低(
P<0.05),LC3B,p62蛋白表达量明显增加(
P<0.01),符合BafA1自噬抑制剂蛋白表达变化规律
[13]。与BafA1组相比,经APS干预后LC3B蛋白表达降低(
P<0.05),符合APS能减少自噬小体合成的推论
[14]。
综上结果显示,APS能增强细胞活力以及增殖能力,APS能抑制自噬的发生,并且APS对RAPA的促自噬作用有拮抗效果,可能与APS能够下调LC3B、Becline-1蛋白表达量有关。
3 讨论
自噬是一种防御和应激调控机制,在缺血性脑卒中可以造成血管内皮细胞不可逆的损伤或死亡
[15],在大鼠脑缺血再灌注损伤后自噬激活,24 h后达到峰值,且此时脑损伤最严重
[16]。林超等研究采用动脉环模型和内皮细胞模型,降低自噬的水平可有效恢复血管生成的能力
[17]。
在本研究中构建体外血管新生模型中,本研究根据国外 Zhu WH 团队研究的血管新生模型的时间点进行实验研究
[11],实验结果显示,体外三维培养模型制备新生血管的时间点为6 d,与该团队的结果一致。结果表明,与对照组比较,6 d后APS组给予黄芪多糖,稳定存在的新生血管数量增多。
为进一步确定机制,本实验在细胞水平进行研究。CCK-8结果显示,在缺氧条件下,与模型组比较,APS作用于内皮细胞后,内皮细胞增殖活性明显增强,结果与朱潇潇团队研究结果一致
[18]。与模型组比较,RAPA作用后的内皮细胞增殖活性明显减弱;BafA1作为抑自噬阳性药,与模型组比较,与APS效果一致,作用于内皮细胞后增强了其增殖活性,但与BafA1组相比,使用APS与BafA1共同干预,内皮细胞增殖活性无明显差别。与RAPA组比,APS与RAPA共同作用复使细胞增殖活性增强(
P<0.01)。此结果说明APS促进RBMEC增殖可能与自噬有关。
Becline-1、LC3B是自噬发生的关键调控分子,当自噬发生时,Becline-1、LC3B的蛋白表达会增加。p62为自噬底物,当自噬被激活时自噬体与溶酶体融合,自噬囊泡中p62等蛋白或细胞器被溶酶体酶降解,p62水平降低
[19],即自噬激活时,p62蛋白表达降低。实验结果中,对比模型组,BafA1提高了LC3B自噬蛋白的表达,原因可能为BafA1为自噬晚期抑制剂,使用BafA1后,细胞内自噬溶酶体降解被抑制,产生自噬体的堆积
[20]。
在本研究中,Western blot 结果表明,在缺氧条件下,与模型组比较,APS可降低Becline-1、LC3B的表达;RAPA可升高Becline-1、LC3B的表达,降低p62的表达。与RAPA组相比,在APS与RAPA共同作用下,Becline-1、LC3B的表达降低,p62表达升高,这提示APS或对RAPA的促自噬作用产生拮抗效果。其中p62表达升高可能与APS能够抑制自噬小体合成有关,但是在单独使用APS干预的组别中,对比模型组,p62蛋白的表达没有明显变化,这可能需要进一步探讨。文中与BafA1组相比,使用APS与BafA1共同干预缺氧的内皮细胞,发现Becline-1表达显著增高,可能的原因为在APS与BafA1的共同干预下,细胞被过度抑制自噬,自噬小体堆积,或促使了自噬的进一步的发生,或导致细胞凋亡,最终表现为Becline-1蛋白表达的增高。
综上所述,我们选用了体外血管段培养模型,细胞的缺氧模型,在细胞基础上分别给予药APS、RAPA、BafA1、APS+RAPA、APS+BafA1进行干预。结果显示,缺氧条件下,APS抑制Becline-1、LC3B的蛋白表达,促进内皮细胞增殖活性,促进血管新生的发生。但其具体的作用方式,自噬是否为促进血管新生的直接因素,仍需要进一步的研究和讨论。
4 结论
本研究基于自噬在缺氧条件下黄芪多糖促进血管新生的作用与机制,结果显示,黄芪多糖能够抑制自噬促进血管内皮细胞增殖,促进新生血管生成。说明黄芪多糖对缺血性脑卒中有一定的治疗作用,其作用机制可能是通过自噬途径实现的。但由于自噬作用机制复杂,对于更深入的作用机制有待于进一步探究。



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}